FENOMENE DE TRANSPORT MEMBRANAR
1. Structura membranei celulare
2. Mecanisme de transport membranar
3. Mecanisme de macrotransport
4. Receptori si traductori de informatie in membranele biologice
1. Structura membranei celulare
Marea majoritate a lipidelor continute de diverse membrane celulare sunt gliceride; ele deriva din molecula de glicerol, care prezinta trei grupari hidroxil (OH). In cazul celulelor eucariote, doua dintre aceste grupari OH sunt legate fiecare la cate un lant de acizi grasi (lant hidrocarburic), printr-o reactie de condensare care duce la formarea unei legaturi esterice intre glicerol si acidul gras respectiv. La fosfolipide, al treilea hidroxil al glicerolului se leaga la o grupare fosfat (PO4ˉ) in urma unei reactii de condensare intre glicerol si acidul fosforic (H3PO4).
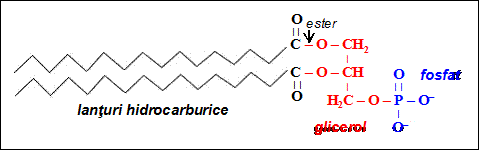
Gruparea fosfat se poate lega in continuare la una din urmatoarele patru grupari polare: colina, etanolamina, serina si inositol, rezultand cele patru tipuri de fosfolipide: fosfatidilcolina (lecitina), fosfatidiletanolamina, fosfatidilserina si fosfatidilinositol.
Toate aceste modificari induc structura specifica fosfolipidelor, care au un capat polar, hidrofil si doua lanturi hidrocarburice nepolare, hidrofobe. Fosfolipidele sunt de aceea molecule amfifile (prezinta atat caracter hidrofil cat si caracter hidrofob). Capatul polar al fosfolipidelor are o sarcina electrica neta negativa.
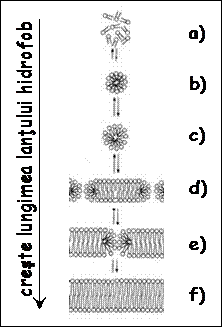 Intr-o solutie apoasa, fosfolipidele formeaza de la sine
structuri complexe cum ar fi micelele
si bistraturile, in care
lanturile hidrofobe se dispun spre interior, iar capetele hidrofile spre
exterior, intr-un aranjament care in acest fel minimizeaza energia
libera a sistemului apa lipide. Lipidele care au lanturi
hidrocarburice scurte, continand 2 pana la 4 atomi de carbon, sunt
solubile in apa (a). Pe
masura ce lanturile hidrofobe devin mai lungi, lipidele incep
sa se autoorganizeze spontan in micele
(b-c). Atunci cand lipide 959c21j le au in jur de 10 atomi de carbon in
lanturile hidrocarburice, ele formeaza bistraturi semi-stabile, in care pot ramane capturate molecule
de apa (d-e). Stabilitatea bistratului lipidic creste odata cu
lungimea lanturilor hidrocarburice, in timp ce permeabilitatea bistratului
scade (f). In general, lanturile
lipidelor aflate in componenta membranelor celulare contin 16 20
atomi de carbon.
Intr-o solutie apoasa, fosfolipidele formeaza de la sine
structuri complexe cum ar fi micelele
si bistraturile, in care
lanturile hidrofobe se dispun spre interior, iar capetele hidrofile spre
exterior, intr-un aranjament care in acest fel minimizeaza energia
libera a sistemului apa lipide. Lipidele care au lanturi
hidrocarburice scurte, continand 2 pana la 4 atomi de carbon, sunt
solubile in apa (a). Pe
masura ce lanturile hidrofobe devin mai lungi, lipidele incep
sa se autoorganizeze spontan in micele
(b-c). Atunci cand lipide 959c21j le au in jur de 10 atomi de carbon in
lanturile hidrocarburice, ele formeaza bistraturi semi-stabile, in care pot ramane capturate molecule
de apa (d-e). Stabilitatea bistratului lipidic creste odata cu
lungimea lanturilor hidrocarburice, in timp ce permeabilitatea bistratului
scade (f). In general, lanturile
lipidelor aflate in componenta membranelor celulare contin 16 20
atomi de carbon.
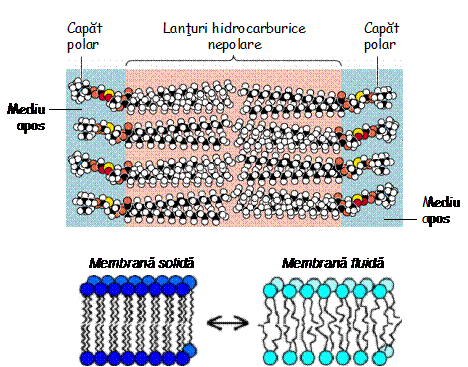
Odata
format, bistratul lipidic este stabilizat prin actiunea unor forte de
slaba intensitate ce determina formarea unor legaturi
necovalente. Forte de tip Van der Waals apar in urma apropierii
spatiale a lanturilor de acizi grasi, determinata de
impachetarea compacta a lipidelor din bistrat. De asemenea, apar
legaturi de hidrogen si atractii electrostatice (legaturi
ionice) intre gruparile hidrofile ale fosfolipidelor si moleculele
solutiei apoase. Spunem ca repulsia hidrofoba determina
formarea bistratului lipidic, iar celelalte forte slabe il
stabilizeaza. Structura bistratului se schimba odata cu
modificarea temperaturii. La temperaturi scazute, lanturile
hidrocarburice ale lipidelor sunt strans impachetate si membrana devine
rigida (se solidifica). Odata cu cresterea temperaturii, lanturile
lipidice devin din ce in ce mai dezordonate, fluiditatea creste, iar
membrana incepe sa se topeasca.
Daca o lipida este plasata intr-o solutie apoasa, moleculele de apa din jurul lipidei nu vor putea forma legaturi de hidrogen cu aceasta. De aceea, pentru a minimiza energia libera a sistemului, moleculele de apa vor tinde sa formeze cat mai multe legaturi intre ele si vor avea mai putina libertate de miscare, ceea ce duce la ordonarea moleculelor de apa intr-o structura de tip cristalin, numita clatrat. Un clatrat are in general forma unui poliedru cu 12 sau 16 fete pentagonale, in centrul caruia se afla molecula hidrofoba. In schimb, in cazul in care lipidele sunt suficient de numeroase pentru a se produce separarea celor doua faze (apa/lipide) prin formarea unor structuri lipidice compacte (micele, bistraturi), realizarea clatratilor nu va mai fi posibila si moleculele de apa vor deveni mai dezordonate.
Prin structura sa, bistratul lipidic reprezinta deci o bariera pentru moleculele hidrofile, cum ar fi proteinele, zaharurile, acizii nucleici, diferiti ioni sau molecule polare. In prezent este unanim acceptat modelul mozaicului fluid proteolipidic al membranei, propus de Nicholson si Singer in 1972. Intr-adevar, diverse studii efectuate cu ajutorul microscopului electronic au aratat ca membranele celulare sunt bistraturi lipidice in care sunt incorporate integral sau partial proteine, glicoproteine si lipoproteine.
Proteinele membranare pot fi:
a) proteine intrinseci (sau integrale);
b) proteine extrinseci (sau periferice).
Proteinele integrale sunt amfifile si reprezinta o proportie de 70% din proteinele membranare. Aceste proteine au un caracter predominant hidrofob, datorat reziduurilor nepolare ale proteinei (ai aminoacizilor hidrofobi). De aceea, proteinele intrinseci sunt inglobate in bistratul lipidic sau, de cele mai multe ori, strabat membrana de la o fata la alta. In multe cazuri, ele prezinta portiuni elicoidale (alfa-helixuri) multiple care traverseaza membrana de mai multe ori. Regiunile hidrofile ale unei astfel de proteine raman expuse la solutia apoasa de pe o parte sau alta a membranei. Proteinele intrinseci sunt puternic legate de membrana, avand un rol important in mentinerea stabilitatii membranelor.
Proteinele periferice nu au regiuni hidrofobe, ele asociindu-se frecvent cu diverse proteine integrale sau cu regiuni hidrofile de la suprafata membranei. De aceea, proteinele extrinseci sunt legate relativ slab de membrana.
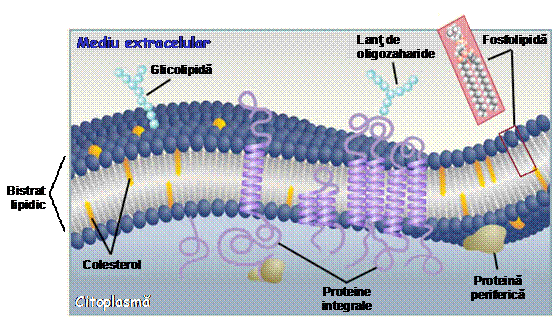
Circa 10% din fosfolipidele si proteinele membranare leaga chimic (prin legaturi covalente) lanturi de oligozaharide (carbohidrati formati din 3-6 monozaharide cum ar fi glucoza, galactoza sau acidul sialic), formand astfel glicolipide si respectiv glicoproteine. Fata exterioara a membranei plasmatice este bogata in glicolipide, ale caror lanturi hidrofobe sunt incorporate in regiunea hidrofoba a membranei, iar capetele hidrofile sunt expuse mediului extracelular. Acestea, impreuna cu carbohidratii atasati la unele proteine integrale, participa la procesele de recunoastere celulara controlate de sistemul imunitar al organismului. Oligozaharidele sunt complet absente pe suprafata interna a membranei plasmatice, ceea ce arata ca tranzitiile rotationale (de pe o fata pe alta a membranei) ale glicoproteinelor si glicolipidelor nu sunt favorizate din punct de vedere energetic. Carbohidratii pot contribui la aranjamentul spatial al glicoproteinelor si pot creste stabilitatea acestora in interiorul membranei, contribuind la rezistenta structurala a membranei. In general, glicoproteinele cresc vascozitatea membranara. In cazul glicolipidelor, oligozaharidele pot actiona ca situsuri de recunoastere pentru anumite molecule din mediul extracelular, pot contribui la stabilitatea membranei sau pot participa la conexiunile dintre celulele vecine intr-un tesut.
Colesterolul este o alta componenta importanta a membranelor celulare, avand un caracter puternic hidrofob. Este un sterol (acesta este o lipida ne-glicerida, o combinatie intre un steroid si un alcool) ce se gaseste in componenta membranelor celulare, in toate tesuturile organismului. Gruparea OH (care este polara) de la capatul moleculei de colesterol interactioneaza cu gruparea fosfat hidrofila a fosfolipidelor membranare, iar portiunea steroida si lantul hidrocarburic nepolar al colesterolului se dispun in interiorul hidrofob al membranei. Colesterolul modifica fluiditatea membranei, stabilizand-o pe intervale mai mari de temperatura; membrana ramane fluida la temperaturi mai scazute decat in absenta colesterolului. Deci, colesterolul scade temperatura de topire a membranelor celulare. De asemenea, colesterolul reduce permeabilitatea membranei la ionii de hidrogen (H+) si de sodiu (Na+).
Fluiditatea membranara este esentiala pentru functionarea corecta a proteinelor integrale. Ea este determinata in principal de compozitia lipidica a membranei si se modifica in functie de proportia lipidelor ale caror lanturi de acizi grasi sunt saturate sau nesaturate. In lanturile hidrocarburice saturate, atomii de carboni sunt legati numai prin legaturi simple C C. Un astfel de lant este flexibil si tinde sa fie liniar. In lanturile hidrocarburice nesaturate unii atomi de carbon pot fi legati prin legaturi duble C = C. Daca lipida este dehidrogenata, prin eliminarea atomilor H se formeaza legaturi de tipul C = C , care determina rasuciri ale moleculei in jurul acestor legaturi. De aceea, lanturile hidrocarburice nesaturate nu se pot impacheta atat de ordonat precum cele saturate. Ca urmare, prezenta unor legaturi de tip C = C in lanturile lipidice de acizi grasi poate duce la coborarea substantiala a temperaturii de topire a lipidelor respective (cu cateva zeci de grade Celsius).
Membrana celulara este o structura dinamica, similara unui fluid vascos, in care moleculele pot executa miscari de translatie si rotatie, cum ar fi:
- translatie in stratul in care se afla (difuzie laterala);
- rotatie in jurul propriei axe;
- flexie (indoire);
- basculare dintr-un monostrat in celalalt (tranzitii rotationale).
Tranzitiile rotationale sunt foarte rare; proteinele integrale in special nu efectueaza astfel de tranzitii.
La temperatura normala a organismului, vascozitatea unei membrane celulare este de cca. 100 de ori mai mare decat cea a apei, astfel incat o fosfolipida se deplaseaza cu 1 μm in interiorul membranei in cca. 1 minut.
2. Mecanisme de transport membranar
Cel mai important rol al unei membrane celulare este de a mentine o frontiera permanenta intre mediul intern si mediul exterior structurii celulare respective. In acest fel compozitia mediului intern este strict controlata de permeabilitatea selectiva a membranei si de diverse procese de transport prin membrana. Ca un caz particular, prezenta in sange a fosfolipazelor (enzime care degradeaza fosfolipidele) poate induce hemoliza pierderea integritatii structurale a membranei plasmatice eritrocitare, ceea ce determina varsarea continutului hematiilor in sange.
Bistratul lipidic ofera o bariera energetica foarte inalta pentru moleculele de dimensiuni mari, ca si pentru moleculele mici, hidrofile. In schimb, bariera energetica este joasa pentru moleculele hidrofobe aflate in mediul apos si cu cat o molecula este mai hidrofoba, cu atat ea va intra mai usor in membrana. Totusi, odata intrata in regiunea hidrofoba a membranei, o molecula nepolara va avea de trecut o bariera energetica relativ inalta pentru a putea parasi membrana.
Capacitatea membranelor celulare de a permite unor molecule hidrofile sa traverseze membrana se datoreaza atat existentei unor proteine membranare integrale (care fie pot forma pori in interiorul membranei, fie transporta ele insele molecule hidrofile de pe o parte pe alta a membranei), cat si permeabilitatii intrinseci a membranei fata de anumite molecule. In general, un bistrat lipidic intact este permeabil la molecule hidrofobe (inclusiv gaze: O2, CO2) si molecule polare mici, neionizate (de exemplu H2O) si nu este permeabil fata de molecule polare de dimensiuni mari (de exemplu zaharuri) si ioni (indiferent de marimea lor).
Transportul diverselor molecule prin membrana celulara se poate realiza fie cu consum de energie celulara metabolica (transport activ) fie fara consum de energie metabolica (transport pasiv).
TRANSPORTUL PASIV
In procesele de transport pasiv moleculele transportate se deplaseaza prin miscari de agitatie termica precum si prin miscari determinate de fortele de atractie sau respingere electrostatica intre moleculele respective si moleculele membranei sau ale mediilor apoase; ca urmare, moleculele transportate se deplaseaza in sensul gradientului potentialului lor electrochimic (daca potentialul electric este acelasi pe ambele fete ale membranei, moleculele se vor deplasa in sensul gradientului de concentratie).
Prin transport pasiv sistemul are tendinta de a ajunge la echilibru termodinamic. Daca in expresia diferentei de potential electrochimic notam c1 = cin, c2 = cex, V1 = Vin, V2 = Vex obtinem:
DW = Win - Wex = RT ln cin/cex + zF (Vin - Vex)
Daca DW > 0 - ionii au tendinta de a parasi celula.
Daca DW < 0 - ionii au tendinta de a patrunde in celula, daca membrana este permeabila pentru acestia. Transportul unei specii ionice inceteaza la echilibru, cand DW = 0. In acest caz potentialul de membrana, E, satisface ecuatia Nernst :
E = Vin - Vex = (RT/zF) ln cex/cin
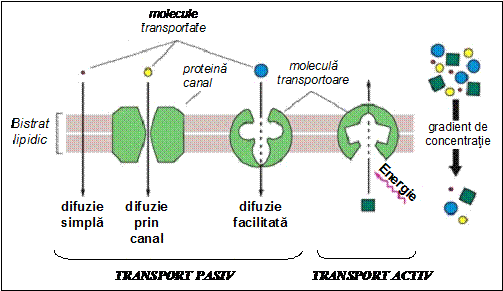
Exista trei mecanisme distincte de transport pasiv:
- difuzie simpla prin dizolvare in bistratul lipidic;
- difuzie simpla prin canale;
- difuzie facilitata.
Difuzia simpla a moleculelor prin dizolvare in bistratul lipidic se datoreaza permeabilitatii intinseci a acestuia fata de anumite tipuri de molecule si ioni (discutate mai sus) si consta in traversarea bistratului fara ajutorul proteinelor membranare.
 Prin
agitatie termica, molecula transportata poate depasi
barierele energetice impuse de diversele regiuni hidrofile/hidrofobe ale
membranei. Deoarece insa ciocnirile moleculare sunt aleatoare, traiectoria
moleculei prezinta numeroase deflexii; molecula poate fie sa
traverseze membrana, fie sa se intoarca in mediul initial.
Transportul unei singure molecule nu depinde de sensul gradientului
potentialului electrochimic. Fluxul net de substanta care
traverseaza membrana prin difuzie simpla are insa sensul
gradientului de potential electrochimic, deoarece moleculele care vor
difuza dinspre mediul cu concentratie mai mare spre mediul cu
concentratie mai mica vor fi mai numeroase decat cele care
traverseaza membrana in sens invers. Schimbul gazos la nivelul alveolelor
pulmonare este un exemplu de transport pasiv realizat prin difuzia simpla
a moleculelor de CO2 si O2 prin dizolvare in
membrana.
Prin
agitatie termica, molecula transportata poate depasi
barierele energetice impuse de diversele regiuni hidrofile/hidrofobe ale
membranei. Deoarece insa ciocnirile moleculare sunt aleatoare, traiectoria
moleculei prezinta numeroase deflexii; molecula poate fie sa
traverseze membrana, fie sa se intoarca in mediul initial.
Transportul unei singure molecule nu depinde de sensul gradientului
potentialului electrochimic. Fluxul net de substanta care
traverseaza membrana prin difuzie simpla are insa sensul
gradientului de potential electrochimic, deoarece moleculele care vor
difuza dinspre mediul cu concentratie mai mare spre mediul cu
concentratie mai mica vor fi mai numeroase decat cele care
traverseaza membrana in sens invers. Schimbul gazos la nivelul alveolelor
pulmonare este un exemplu de transport pasiv realizat prin difuzia simpla
a moleculelor de CO2 si O2 prin dizolvare in
membrana.
Difuzia simpla prin canale ionice sau moleculare permite un transport molecular extrem de rapid prin membrana. Canalele ionice joaca de aceea un rol extrem de important in procesele de semnalizare celulara, in care viteza de transmitere a informatiei este un element critic. Un canal membranar este o proteina integrala complexa care formeaza o cale de trecere (un por) pentru anumite molecule sau ioni insolubili in matricea lipidica a membranei.
Activitatea unui canal ionic sau molecular este discontinua: perioadele in care canalul este deschis alterneaza cu perioade de inactivitate, in care canalul este inchis. Prin diverse moduri de reglare a activitatii canalelor membranare, celula controleaza fluxurile de substanta prin membrana. In plus, canalele membranare prezinta o mare specificitate pentru anumiti ioni sau molecule, nepermitand altor tipuri de ioni sau molecule sa le traverseze.
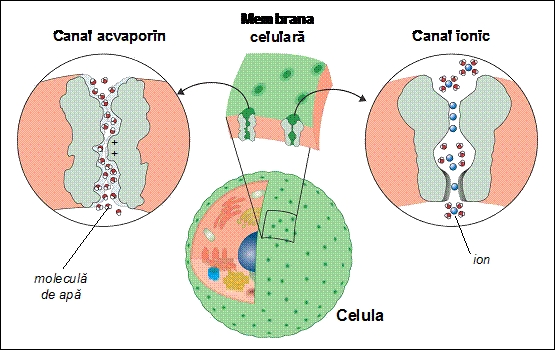
Aminoacizii hidrofobi ai unei proteine formatoare de pori se orienteaza spre regiunea lipidica hidrofoba a membranei, in timp ce aminoacizii hidrofili ai proteinei se orienteaza spre interiorul porului, pentru a putea interactiona cu molecula hidrofila care traverseaza membrana prin por. De exemplu, canalul acvaporin (AQP) permite numai trecerea moleculelor de apa, care se orienteaza in campul electric produs de atomii aflati in peretele interior al canalului. In schimb, ionii pozitivi (cum ar fi H+, H3O+, Na+) sunt respinsi de sarcinile pozitive aflate in regiunea centrala a porului. Exista deci in structura canalului o anumita regiune, numita filtru, care determina proprietatile selective ale canalului. In general, canalele ionice (de exemplu canalele ionice de sodiu, potasiu, calciu, clor) au selectivitate foarte mare pentru un anumit tip de ioni (Na+, K+, Ca2+ si respectiv Cl- in exemplul anterior), dar pot permite in acelasi timp trecerea altor tipuri de ioni, desi in numar mult mai redus. Un caz particular este reprezentat de canalele de potasiu care permit trecerea ionilor K+ dar nu si a ionilor Na+, desi ionii de sodiu sunt mai mici decat cei de potasiu si au aceeasi sarcina electrica. Mecanismul de filtrare al acestui canal de K+ se bazeaza pe interactia dintre ionul transportat si atomii de oxigen aflati in structura filtrului. In mediul apos din exteriorul canalului, atat ionii Na+ cat si ionii K+ sunt legati la cate 4 molecule de apa. La intrarea in filtru, ionii se desprind de moleculele de apa si se leaga la atomii de oxigen ai filtrului. Pentru atomul K+, distanta de interactie cu 4 molecule de oxigen ale filtrului este egala cu distanta de legatura cu cele 4 molecule de apa din mediul apos, in timp ce pentru ionul Na+ aceasta distanta este mai mica. Pentru a putea traversa regiunea corespunzatoare filtrului, ionul trebuie sa culiseze de-a lungul canalului prin legari succesive la cate 4 atomi de oxigen simultan, ceea ce nu este posibil decat in cazul ionului de potasiu. Descoperirea acestui mecanism de filtrare selectiva a ionilor pentru canalul de potasiu a fost realizata prin cristalografie cu raze X, pentru care Roderick MacKinnon a primit Premiul Nobel in anul 2003.
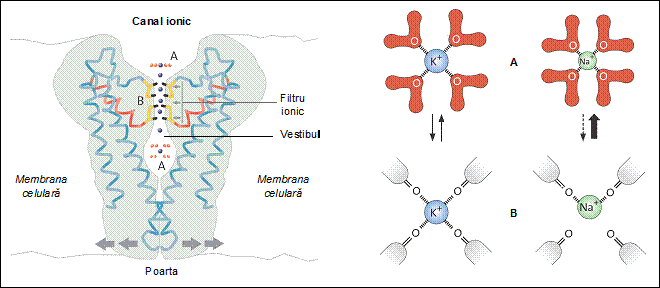
In structura
unui canal ionic se disting cateva domenii caracteristice: filtrul ionic, vestibulul,
poarta si senzorul. In vestibul ajung numai ionii selectati in filtru;
filtrul nu permite trecerea moleculelor mari, cum ar fi de exemplu glucoza.
Poarta se inchide sau se deschide in functie de reactia senzorului la
diferitele semnale receptionate. Senzorul poate fi o regiune externa
a proteinei-canal, aflata in contact cu mediul apos de pe una din
fetele membranei, sau o regiune interna a proteinei. Semnalele la care este sensibil senzorul
pot fi chimice (anumite molecule din
mediul apos care se leaga la situsuri specifice ale proteinei-canal), electrice (diferenta de
potential electric intre cele doua medii apoase pe care le
separa membrana, denumita si potential transmembranar sau
potential de membrana), mecanice
(presiunea), termice (caldura)
sau electromagnetice (lumina).
Difuzia facilitata este transportul de substanta prin membrana fie prin intermediul unor proteine transportoare liposolubile, care pot sa traverseze membrana prin dizolvare (transportori difuzibili), fie prin intermediul unor proteine membranare integrale, care trec molecula hidrofila de pe o parte pe alta a membranei printr-o modificare in propria conformatie sau prin formare de pori in membrana. Moleculele transportoare (care se mai numesc transportori sau ionofori) nu au proprietati enzimatice; in procesul de difuzie facilitata nu se consuma energie; deplasarea moleculei transportate se face in sensul gradientului sau electrochimic. Exista o multitudine de molecule transportoare care asigura difuzia facilitata a multor tipuri de ioni sau molecule (de exemplu glucoza, colina, ionii K+). Majoritatea moleculelor transportate in acest mod nu pot trece nici prin canale ionice (datorita marimii lor), nici prin matricea lipidica (deoarece nu sunt liposolubile). In numeroase celule, mecanismul principal prin care glucoza din mediul extracelular intra in citoplasma este difuzia facilitata. Deoarece exista un numar limitat de molecule care pot transporta glucoza prin membrana plasmatica, influxul de glucoza nu poate depasi o anumita limita nici chiar atunci cand concentratia extracelulara de glucoza este foarte mare.
Un exemplu de ionofor este antibioticul valinomicina, care se insereaza in membranele celulare, unde se comporta ca o proteina intrinseca. Atunci cand este incorporata in membrana, o molecula de valinomicina creste viteza de transport prin membrana a ionilor K+ la 104 ioni/s.
Un exemplu de
ionofor formator de por este un alt antibiotic, gramicidina A. Moleculele de
gramicidina patrund in interiorul bistratului lipidic, unde
formeaza dimeri helicoidali care strabat membrana, determinand astfel
aparitia unor pori conductori cilindrici. Fiind o proteina formatoare
de pori, gramicidina isi expune gruparile hidrofobe spre zona
lipidica a membranei, in timp ce gruparile hidrofile sunt orientate
spre interiorul porului. Acesta nu permite trecerea decat a ionilor Na+;
fluxul de difuzie pentru o molecula de antibiotic este de 107
ioni/s. Tot ionofori formatori de pori sunt si alametacina si
monazomicina.
Intre difuzia facilitata si difuzia prin canale exista cateva deosebiri importante:
- transportorii au o specificitate mai mare; acestia pot distinge intre speciile levogire si dextrogire, astfel incat in celula pot patrunde prin difuzie facilitata numai glucoza dextrogira si aminoacizii levogiri. Pot transporta mii de molecule/s.
- canalele au o viteza mult mai mare de transport al ionilor (milioane sau chiar sute de milioane de ioni/s). Atunci cand au loc procese care necesita modificari foarte rapide ale concentratiei sau compozitiei ionice (de exemplu in producerea influxului nervos) canalele ionice sunt mai adecvate pentru transportul speciilor implicate in aceste procese.
- transportorii se gasesc in numar mult mai mare decat canalele;
- transportorii pot participa la transportul activ secundar.
TRANSPORTUL ACTIV
Exista doua tipuri de transport activ: primar si secundar.
In procesele de transport activ primar sunt implicate proteine membranare integrale cu proprietati enzimatice, care transporta ioni sau molecule impotriva gradientului lor de concentratie, de potential electric sau de presiune osmotica, adica in sens opus tendintei naturale de crestere a entropiei care sa duca la anularea acestor gradienti. Astfel de proteine se numesc pompe membranare. Transportul activ primar este posibil numai prin cuplarea sa energetica cu reactii care furnizeaza energie libera (reactii exergonice); este de aceea un proces endergonic (consumator de energie). Reactiile exergonice care furnizeaza energia necesara functionarii pompelor membranare sunt in general reactii metabolice; exista insa si pompe care utilizeaza energia luminii.
In membranele celulare exista mai multe tipuri de pompe ionice. Majoritatea pompelor functioneaza pe principiul utilizarii energiei moleculei de ATP pentru a transporta anumiti ioni impotriva gradientului electrochimic intre cele doua fete ale membranei; molecula de ATP se leaga la molecula pompei ionice si este hidrolizata la ADP, proces in care este eliberata o parte din energia chimica a ATP-ului. Proteina pompei absoarbe partial energia furnizata prin hidroliza ATP si isi schimba conformatia. Prin aceasta modificare conformationala, pompa efectueaza trecerea moleculei hidrofile de pe o parte pe alta a membranei. Energia absorbita de proteina este in final cedata prin relaxarea structurii pompei, care revine la conformatia initiala si ciclul poate fi reluat. Energia care nu este absorbita de pompa este disipata prin incalzirea locala a mediului apos in care se produce hidroliza ATP. Pompele care folosesc energia ATP se numesc pompe ATP-azice sau ATP-aze.
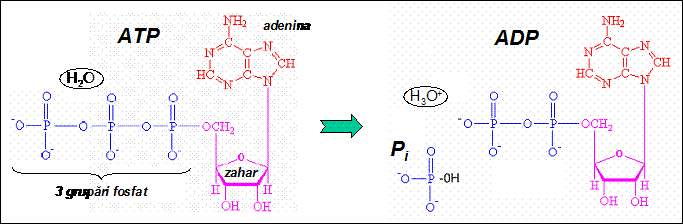
Procesul de transport activ primar este mecanismul principal prin care sunt mentinute diferentele de concentratii ionice intre diferitele timente celulare, precum si intre citosol si mediul extracelular. In cardiomiocite, cel mai important rol in mentinerea potentialului de repaus la valoare constanta il au pompele de Na+-K+ ale plasmalemei, care extrag trei ioni de sodiu din celula si introduc doi ioni de potasiu din mediul extracelular in celula pentru fiecare molecula de ATP hidrolizata. Modul de functionare este urmatorul. In conformatia de repaus, pompa prezinta pe partea intracelulara o afinitate foarte mare pentru ionii Na+ si foarte mica pentru ionii K+. Ca urmare, ionii Na+ ajunsi prin difuzie din mediul intracelular la situsurile de legare ale pompei se leaga la acestea, stimuland hidroliza moleculei de ATP care este legata de pompa. Ca urmare, pompa isi schimba conformatia. In noua conformatie, situsurile de legare ale sodiului sunt expuse mediului extracelular, iar accesul ionilor Na+ dinspre regiunea intracelulara este blocat. Tot in noua conformatie, afinitatea pompei pentru Na+ este foarte mica, astfel incat ionii Na+ se desprind de situsurile pompei si difuzeaza in mediul apos extracelular. In schimb, in aceasta conformatie afinitatea proteinei fata de ionii K+ este foarte mare, ceea ce favorizeaza legarea rapida a ionilor de potasiu la situsurile pompei; aceasta legare induce relaxarea proteinei la conformatia initiala. Ionii K+ legati la proteina sunt acum expusi mediului intracelular. Deoarece afinitatea fata de K+ este din nou redusa, ionii K+ se desprind si difuzeaza in mediul intracelular; gruparea fosfat paraseste proteina, permitand legarea unei alte molecule ATP in locul sau.
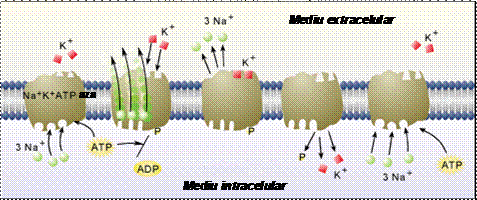
O singura molecula de Na+-K+-ATP-aza scindeaza in jur de 100 molecule de ATP pe secunda, iar la fiecare ciclu de functionare transfera o sarcina electrica neta pozitiva (+e) spre exterior, ceea ce contribuie la polarizarea electrica a membranei. Aceasta proprietate defineste caracterul electrogenic al pompei. Pompa de Na+-K+ se gaseste in plasmalema tuturor celulelor de origine animala. In repaus, cca. 30% din energia ATP-ului celular este consumata pentru functionarea acestei pompe. In celulele nervoase, in faza de repolarizare acest consum ajunge la 70% din consumul energetic celular.
In membranele celulare exista si alte tipuri de pompe ionice. Un rol foarte important il are Ca2+-ATP-aza, care mentine o concentratie foarte redusa de Ca2+ in citosol, de ordinul ~100 nM. Pentru fiecare molecula de ATP hidrolizata, Ca2+-ATP-aza pompeaza cate doi ioni de calciu din citosol. Datorita pompelor de calciu din plasmalema si din membrana reticulului endo/sarcoplasmatic, gradientul ionilor de calciu este mentinut la o valoare foarte mare (3-4 ordine de marime) atat intre citosol si mediul extracelular, cat si intre citosol si interiorul reticulului endo- sau sarcoplasmatic (in conditii normale, concentratia Ca2+ atat in reticul cat si mediul extracelular este in jur de 1 mM). Daca functionarea Ca2+-ATP-azei este deficitara, celula va avea un nivel toxic al calciului citosolic, ceea ce induce apoptoza (moartea celulara programata).
Alte pompe ionice sunt H+-K+-ATP-aza din membrana celulelor mucoasei gastrice sau pompa protonica bacteriorodopsina aflata in membranele unor bacterii. Sub actiunea luminii, care induce o reactie de fotoizomerizare in bacteriorodopsina, aceasta sufera o modificare conformationala prin care transloca un proton din citoplasma in exteriorul celulei.
In general, pompele membranare sunt reversibile. In cazul in care concentratiile de sodiu extracelular si de potasiu intracelular sunt excesiv de mari, iar concentratiile de ADP si fosfat intracelular (Pi) sunt suficient de mari, ciclul pompei de Na+-K+ poate functiona in sens invers, iar aceasta incepe sa sintetizeze ATP. Un alt exemplu remarcabil este ATP-sintaza, care se afla in membrana mitocondriilor, cloroplastelor sau ale bacteriilor aerobe sau fotosintetice. Aceasta enzima complexa fie sintetizeaza ATP folosind gradientul existent de H+, fie functioneaza ca H+-ATP-aza, producand un gradient de protoni. In cele doua cazuri, deplasarea protonilor are loc in sensuri opuse.
In anumite situatii, celulele pot de asemenea sa sintetizeze proteine care sa formeze pompe membranare, pentru a elimina anumite substante toxice sau medicamente. Prin acest mecanism, de exemplu, in chimioterapia cancerului apare rezistenta la medicamentele administrate.
 In procesele de transport activ secundar
sunt implicati transportori complecsi, care cupleaza doi
gradienti electrochimici diferiti, astfel incat energia
eliberata prin miscarea unor molecule de un anumit tip in sensul
gradientului lor electrochimic este folosita pentru a transporta molecule
de alt tip impotriva gradientului lor electrochimic. Astfel de transportori se
numesc cotransportori, iar procesul
se numeste cotransport sau transport cuplat. Exita simport, in care ambele tipuri de
molecule sunt transportate in aceeasi directie, si antiport, prin care cele doua
tipuri de molecule sunt transportate in sensuri opuse. Datorita proceselor
celulare de pompaj ionic, membrana plasmatica prezinta o sarcina
neta negativa pe fata interioara si o sarcina
neta pozitiva pe fata exterioara, ceea ce determina o
valoare negativa a diferentei de potential membranar. In
cotransport, deplasarea unor sarcini electrice in sensul potentialului lor
electrochimic reprezinta un proces exergonic deoarece sistemul tinde in
acest fel la starea de echilibru in care diferenta de potential
membranar sa se anuleze, deci tinde la o stare cu energie mai mica.
Energia cedata in acest proces este preluata de cotransportori pentru
a transloca alte molecule sau ioni impotriva gradientului electrochimic, de
aceea acest tip de transport este activ.
In procesele de transport activ secundar
sunt implicati transportori complecsi, care cupleaza doi
gradienti electrochimici diferiti, astfel incat energia
eliberata prin miscarea unor molecule de un anumit tip in sensul
gradientului lor electrochimic este folosita pentru a transporta molecule
de alt tip impotriva gradientului lor electrochimic. Astfel de transportori se
numesc cotransportori, iar procesul
se numeste cotransport sau transport cuplat. Exita simport, in care ambele tipuri de
molecule sunt transportate in aceeasi directie, si antiport, prin care cele doua
tipuri de molecule sunt transportate in sensuri opuse. Datorita proceselor
celulare de pompaj ionic, membrana plasmatica prezinta o sarcina
neta negativa pe fata interioara si o sarcina
neta pozitiva pe fata exterioara, ceea ce determina o
valoare negativa a diferentei de potential membranar. In
cotransport, deplasarea unor sarcini electrice in sensul potentialului lor
electrochimic reprezinta un proces exergonic deoarece sistemul tinde in
acest fel la starea de echilibru in care diferenta de potential
membranar sa se anuleze, deci tinde la o stare cu energie mai mica.
Energia cedata in acest proces este preluata de cotransportori pentru
a transloca alte molecule sau ioni impotriva gradientului electrochimic, de
aceea acest tip de transport este activ.
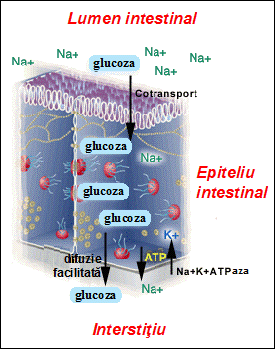
Cotransportul este implicat de exemplu in transferul glucozei din intestin in sange prin celulele epiteliului intestinal. In membrana bazala (dinspre interstitiu) a celulelor epiteliale se gasesc pompe de Na+-K+, care mentin o concentratie scazuta de ioni Na+ in interior (10-l5 mM fata de 144 mM in exterior si 144 mM in lumenul intestinului). In membrana apicala (dinspre lumen) se gasesc cotransportori care transfera simultan (prin simport) cate un ion Na+ si o molecula de glucoza (sau un aminoacid) in citoplasma; in acest proces glucoza este transportata impotriva gradientului de concentratie. In acest caz, cotransportul este electrogenic deoarece are ca rezultat net transportul unei sarcini electrice pozitive dintr-o parte a membranei in cealalta. Concentratia de glucoza in citoplasma este mai mare decat in interstitiu, iar la nivelul membranei bazale glucoza este transportata pasiv prin difuzie facilitata. Efectul net al acestor procese de transport este transferul transcelular de glucoza din spatiul luminal spre sange. Transportul glucozei din lumen in interstitiu este blocat daca pompa ionica de Na+-K+ este inhibata (de exemplu cu oligomicina sau ouabaina).
In celulele nodurilor sinusal si atrioventricular se produce de asemenea un transfer de sarcina prin antiport electrogenic 3 ioni Na+/ 1 ion Ca2+ (influx de sarcina neta +e pentru fiecare molecula cotransportoare). Cotransportorul Na+/Ca2+ transloca deci un ion de calciu din interior in mediul extracelular pe baza energiei eliberate la deplasarea a trei ioni Na+ in sensul gradientului electrochimic, in timp ce Na+-K+-ATP-aza din plasmalema pompeaza ionii Na+ din interior spre exterior.
|
Ionul |
Concentratia extracelulara (mM) |
Concentratia intracelulara (mM) |
|
Na+ |
145 |
15 |
|
K+ |
4 |
150 |
|
Ca2+ |
2 |
10-4 |
Pompele de Ca2+ si
Na+-K+ asigura gradienti mari ai ionilor Ca2+,
Na+ si K+ intre sarcoplasma si mediul
exterior in celula relaxata, precum si revenirea la valorile normale
ale acestora dupa generarea potentialelor electrice (potentiale
de actiune). De asemenea, antiportul Na+/Ca2+ are un
rol esential in generarea periodica spontana a
potentialelor de actiune de catre celulele nodurilor, deoarece
participa la acumularea de sarcini pozitive pe fata interioara a
sarcolemei si deci la depolarizarea progresiva a celulelor.
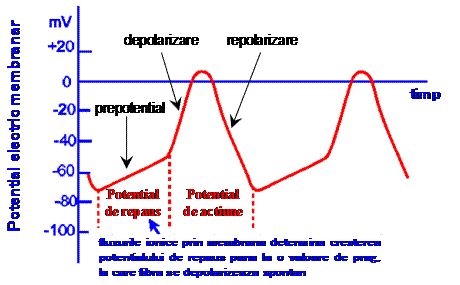
Ca urmare, potentialul electric al membranei creste continuu, de la aproximativ -70 mV pana la valoarea de prag, de aproximativ -50 mV (faza prepotentialului de actiune). Cand potentialul depaseste valoarea de prag, se produce activarea rapida a canalelor de sodiu si calciu din sarcolema, ceea ce determina cresterea influxului de cationi Na+ si Ca2+ in celula. Potentialul electric membranar creste rapid la valori pozitive (faza de depolarizare rapida), care determina inchiderea canalelor de calciu si sodiu. Urmeaza faza de repolarizare, in care excesul de cationi acumulati in sarcoplasma este eliminat prin actiunea pompelor de sodiu si calciu. Concentratiile diferitilor ioni revin lent la valoarea de repaus, ceea ce determina scaderea potentialului membranar la valoarea de repaus.
In procesele de macrotransport substantele intra sau ies din celula sub forma unor aglomerari multimoleculare. In procesele de transport membranar discutate mai sus moleculele sau ionii erau transportati individual prin membrana; de aceea transportul membranar se mai numeste microtransport. Desi in procesele de macrotransport diverse substante sunt introduse sau scoase din celula, aceasta nu se realizeaza prin traversarea propriu-zisa a membranei plasmatice, ci prin formare de cule membranare care sunt inglobate sau eliminate din celula.
Exista trei tipuri de macrotransport: endocitoza, exocitoza si transcitoza.
In procesul de endocitoza materialul extracelular este introdus in interiorul celulei, fara a fi insa dispersat in citoplasma (ramane in continuare ca material extra-citoplasmatic). Cele trei modalitati de macrotransport prin endocitoza sunt: fagocitoza, pinocitoza si endocitoza mediata de receptori.
In fagocitoza celula inglobeaza particule extracelulare solide pe care le inconjoara cu pseudopode (prelungiri citoplasmatice) care apoi fuzioneaza in spatele lor. Se formeaza astfel o vacuola in citoplasma, care contine materialul ingerat. In organismul uman exista celule specializate (fagocite, macrofagi) care prin fagocitoza inglobeaza si distrug diverse substante toxice, reziduuri celulare apoptotice sau bacterii.
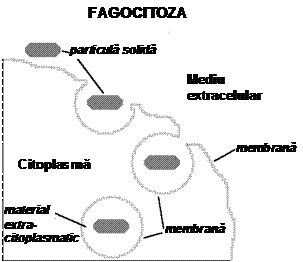
In pinocitoza celula inglobeaza lichid extracelular in mod similar fagocitozei.
In endocitoza mediata de receptori, anumite substante extracelulare se leaga la receptori membranari specifici, care induc formarea de cule membranare in care este inglobat material extracelular. De exemplu, prin acest mecanism celulele organismului preiau colesterolul transportat de sange. De asemenea, virusul gripal intra in celule prin endocitoza mediata de receptori.
Endocitoza prezinta avantajul de a minimiza volumul in care trebuie sa actioneze enzimele de digestie a materialului internalizat. Acestea descompun materialul in molecule mici prin digestie (de obicei prin hidroliza). O parte din moleculele rezultate pot permea membrana culei, trecand in citoplasma. Veziculele rezultate prin endocitoza pot de asemenea sa fuzioneze cu lizozomii, in care are loc degradarea unui material bogat in lipide, proteine si carbohidrati proveniti din membranele celulare.
In procesul de exocitoza, cule membranare continand material intracelular sunt transportate spre membrana plasmatica, fuzioneaza cu aceasta si elibereaza materialul in exteriorul celulei. Prin acest mecanism se produce, de exemplu, secretia de insulina, eliberarea anticorpilor in sange sau eliberarea de neurotransmitatori (acetilcolina, glutamatul etc.) in terminatiile nervoase.
In procesul de transcitoza, cule cu material extracelular sunt inglobate in celula, traverseaza celula fara a-si deversa continutul in citoplasma, apoi fuzioneaza cu membrana plasmatica pe partea opusa a celulei si elibereaza materialul in spatiul extracelular. De exemplu, transcitoza prin endoteliul capilar determina trecerea proteinelor plasmatice din sange in spatiul extravascular.
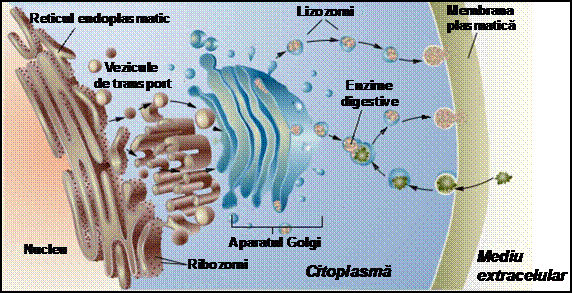
Membranele celulare sunt in mod continuu degradate, reciclate si regenerate. Diversii componenti membranari circula intre diferite timente celulare si suprafata celulei intr-un flux permanent, mentinut prin procese de endocitoza si exocitoza. Componentele membranelor celulare sunt sintetizate in reticulul endoplasmatic, transportate ca cule spre aparatul Golgi unde sunt modificate, iar apoi sunt transportate spre suprafata celulara. Aici culele isi elibereaza continutul in mediul extracelular (prin exocitoza), iar compusii membranari eliberati se insereaza sau se ataseaza la membrana. Degradarea componentelor membranare se produce in lizozomi, sub actiunea unor enzime digestive. Daca una sau mai multe enzime lizozomice prezinta activitate defectuoasa, apar unele afectiuni asociate cu stocarea excesiva de material membranar in lizozomi. De exemplu, in boala Tay-Sachs (boala lizozomala), enzima hexozaminidaza A este defectuoasa, iar glicolipidele nu mai pot fi degradate. Acumularea de glicolipide in lizozomi induce dereglari neurologice grave si moarte. Alte mutatii pot determina de asemenea stocarea proteinelor-receptor in reticulul endoplasmatic. Incapacitatea de a forma receptori in membrana plasmatica duce, de exemplu, la dezvoltarea a numeroase afectiuni ale cordului.
Receptorii membranari sunt proteine integrale complexe care in urma receptiei unui stimul din mediul extracelular (denumit semnal primar sau mesager prim) produc un semnal secundar (denumit mesager secund) care determina o serie de procese celulare (acestea definesc raspunsul celular la stimulul extern). Receptorii sunt capabili sa interactioneze rapid si reversibil cu mesagerii primi, care pot fi mediatori chimici, hormoni, antigeni, medicamente etc. Mesagerul secund poate fi uneori chiar complexul receptor mesager prim, dar in general este o molecula sintetizata de celula, care poate fi:
- hidrofoba, atasata de membrana, de care se poate desprinde (de ex. diacil glicerolul, DAG; inositol trifosfatul, IP3; lipide de tip fosfatidilinositol); difuzeaza in imediata vecinatate, putand activa fie proteine efectoare care sunt atasate la membrana, fie receptori ai unor organite celulare;
- hidrofila, difuzeaza in citosol (ex. acidul adenozin monofosforic ciclic, c-AMP; acidul guanozin monofosforic ciclic, c-GMP; ionii Ca2+);
- gazoasa, cum ar fi oxidul azotic (NO) sau monoxidul de carbon (CO); poate atat sa difuzeze in citosol cat si sa traverseze membranele celulare.
Fiecare receptor recunoaste un anumit tip de semnal, dar poate lega mai multe tipuri de molecule la situsuri diferite. Aceste molecule, care reprezinta liganzii receptorului respectiv, pot afecta rata de producere a mesagerului secund.
Etapele semnalizarii intercelulare in organism
1. Sinteza moleculei semnal
2. Eliberarea moleculei semnal
3. Transportul moleculei semnal catre tinta
4. Detectia semnalului
5. Raspunsul celular
6. Desprinderea moleculei semnal de receptor
Clasificarea semnalelor
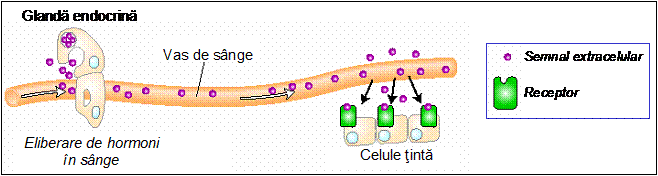
- Semnale endocrine, reprezentate de hormonii produsi in glandele endocrine, secretati in sange si distribuiti in organism;
- Semnale paracrine, generate de celule, actioneaza local in vecinatate. Transmiterea unui semnal (prin intermediul unui neurotransmitator) de la o celula nervoasa la alta sau de la o celula nervoasa la o celula musculara este un exemplu de semnalizare paracrina;
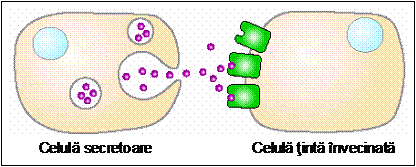
- Semnale autocrine (celulele secreta molecule-semnal care se leaga de receptorii proprii). Celulele mentinute in culturi celulare pot secreta factori de crestere care sa le stimuleze propria proliferare. In general, acest tip de semnalizare este caracteristic celulelor canceroase; acestea produc excesiv factori de crestere care determina o proliferare proprie necontrolata si, in plus, influenteaza celulele netumorale adiacente, putand conduce la formarea unei mase tumorale.
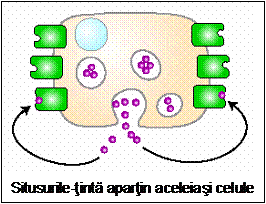
- Semnale de contact - necesita contactul intercelular. Acest tip de semnalizare intervine in special in procesele de diferentiere.
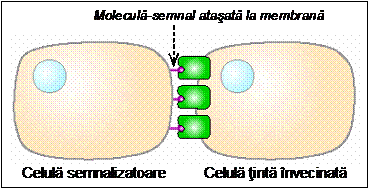
- Semnale nervoase - se transmit de-a lungul axonilor catre celulele tinta.
Clasificarea receptorilor membranari
I. Receptori asociati cu canale ionice (ionotropi)
II. Receptori asociati cu o proteina G
III. Receptori asociati cu enzime (sunt fie ei insisi enzime, fie sunt asociati cu enzime pe care le activeaza). Enzimele respective sunt, in marea lor majoritate, protein kinaze (enzime care pot fosforila alte proteine).
Receptorii din ultimele doua clase mai sunt numiti si receptori metabotropi.
Receptorii ionotropi sunt proteine-receptor care formeaza
canale ionice. Aceste proteine au o zona receptoare
la care se leaga mesagerul prim si o zona efectoare prin care
comunica senzorului canalului comanda de deschidere sau inchidere a
portii.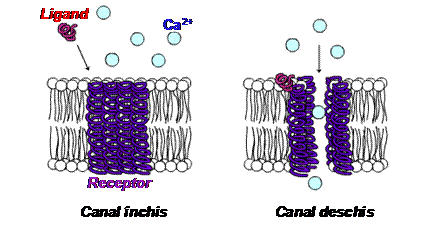
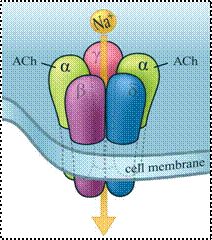
Exemplu: receptorul nicotinic de acetilcolina (nAChR) este format din 5 subunitati similare, dintre care doua subunitati a au fiecare cate un situs de legare pentru moleculele ACh. Prin legarea ACh la subunitatile a ale receptorului, are loc o modificare conformationala in urma careia canalul interior se deschide, permitand trecerea ionilor Na+ in sensul gradientului de potential electrochimic. Receptorul nACh poate fi activat nu numai de acetilcolina, ci si de nicotina, care are un efect mult mai puternic asupra receptorului.
Receptorii asociati cu o proteina G sunt receptori metabotropi care pot determina sinteza mesagerilor secunzi prin intermediul unei proteine efectoare. Acest tip de receptori prezinta 7 domenii (coloane) transmembranare conectate prin bucle intra- si extracelulare. Legarea mesagerului prim la receptor induce o modificare conformationala a proteinei receptorului in urma careia este activata proteina G legata la buclele citoplasmatice II si III ale receptorului. Ca urmare, proteina G se disociaza in subunitatile Ga si Gbg. In forma libera (nelegata), Ga schimba GDP cu GTP, difuzeaza si interactioneaza cu fosfolipaza C (PLC) de tip PLCβ1, care astfel devine activa (capata activitate enzimatica). La randul sau, complexul Gbg activeaza o alta forma a fosfolipazei C, PLCβ2. Cele doua izozime PLC catalizeaza disocierea moleculei de inositol 1,4,5-trifosfat (IP3) de fosfolipida membranara fosfatidilinositol 4,5-difosfat (PIP2), care devine diacilglicerol (DAG); DAG ramane incorporat in membrana. In continuare, IP3 difuzeaza in citosol, determinand activarea canalelor de Ca2+ ale reticulului endoplasmatic (care sunt receptori de IP3), iar DAG activeaza proteinkinaza C (PKC). Calciul eliberat din reticulul endoplasmatic poate actiona si el ca mesager secund pentru alte procese celulare. Atat PKC cat si Ca2+ sunt implicati in numeroase procese celulare. De asemenea, forma activa a PLCβ1 poate functiona ca o proteina de activare a GTP-azei, avand un rol important in proliferarea celulara.
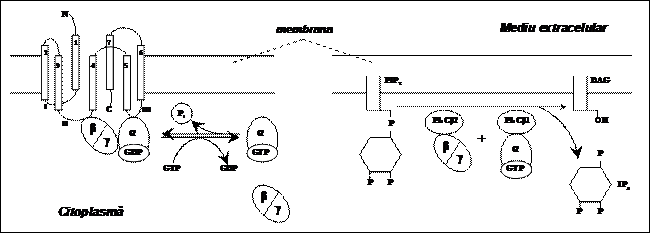
Complexul Ga - GTP poate de asemenea interactiona cu adenilat ciclaza (proteina efectoare), care se activeaza si catalizeaza transformarea ATP din celula in c-AMP (mesager secundar); c-AMP declanseaza o serie de reactii enzimatice prin care este activata enzima fosforilaza. Aceasta determina transformarea glicogenului in glucoza (raspunsul celular). Dupa activarea efectorului, subunitatea a a proteinei G devine inactiva prin hidroliza GTP si se recombina cu subunitatile b si g, refacand proteina G. Procesul se poate relua. Pe acest principiu functioneaza de exemplu receptorul de epinefrina (adrenalina).
Receptorii asociati cu enzime activeaza o protein-kinaza citoplasmatica. De exemplu, receptorul de insulina poate activa o tirozin-kinaza, care fosforileaza alte proteine.
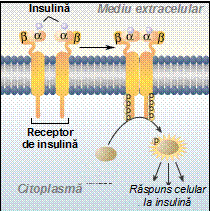 Atunci cand
cele doua subunitati α ale receptorului leaga fiecare
cate o molecula de insulina, proteina receptorului sufera o
modificare in structura, expunandu-si regiunile catalitice spre
citoplasma. In acest fel proteina devine o kinaza activa, care
in continuare fosforileaza alte proteine intracelulare, producand
raspunsul celular (cresterea permeabilitatii pentru
glucoza). Prin acest mecanism, insulina produce un raspuns in
interiorul celulei fara a trece prin bistratul lipidic. De exemplu,
in celulele musculare, activarea receptorului de insulina determina o
serie de reactii enzimatice care conduc la fuzionarea cu membrana
plasmatica a unor cule din citoplasma ce contin proteina
Glut4. Ca urmare, Glut4, care este un transportor al glucozei, se
insereaza in membrana exterioara a celulei, stabilind un influx de
glucoza din mediul extracelular in celula (prin difuzie facilitata).
Atat in muschi cat si in ficat, insulina promoveaza influxul de
glucoza in celule; apoi acestea sintetizeaza glicogen, sursa
energetica celulara in care este inmagazinata energia glucozei.
In plus, ficatul foloseste excesul de glucoza pentru a sintetiza
acizi grasi, care sunt impachetati in granule numite lipoproteine,
eliberate apoi in sange. In celulele tesutului adipos, influxul de
glucoza generat de insulina contribuie la sinteza glicerolului, iar
glucoza in exces induce formarea de triacilgliceride (grasimi), plecand de
la acizii grasi din lipoproteine. Efectul net al insulinei este acumularea
de grasimi in tesutul adipos (daca exista exces de
glucoza).
Atunci cand
cele doua subunitati α ale receptorului leaga fiecare
cate o molecula de insulina, proteina receptorului sufera o
modificare in structura, expunandu-si regiunile catalitice spre
citoplasma. In acest fel proteina devine o kinaza activa, care
in continuare fosforileaza alte proteine intracelulare, producand
raspunsul celular (cresterea permeabilitatii pentru
glucoza). Prin acest mecanism, insulina produce un raspuns in
interiorul celulei fara a trece prin bistratul lipidic. De exemplu,
in celulele musculare, activarea receptorului de insulina determina o
serie de reactii enzimatice care conduc la fuzionarea cu membrana
plasmatica a unor cule din citoplasma ce contin proteina
Glut4. Ca urmare, Glut4, care este un transportor al glucozei, se
insereaza in membrana exterioara a celulei, stabilind un influx de
glucoza din mediul extracelular in celula (prin difuzie facilitata).
Atat in muschi cat si in ficat, insulina promoveaza influxul de
glucoza in celule; apoi acestea sintetizeaza glicogen, sursa
energetica celulara in care este inmagazinata energia glucozei.
In plus, ficatul foloseste excesul de glucoza pentru a sintetiza
acizi grasi, care sunt impachetati in granule numite lipoproteine,
eliberate apoi in sange. In celulele tesutului adipos, influxul de
glucoza generat de insulina contribuie la sinteza glicerolului, iar
glucoza in exces induce formarea de triacilgliceride (grasimi), plecand de
la acizii grasi din lipoproteine. Efectul net al insulinei este acumularea
de grasimi in tesutul adipos (daca exista exces de
glucoza).
Intrebari:
1. Explicati procesul de solidificare a membranelor lipidice la temperaturi scazute.
2. In apa, lipidele pot forma micele sau bistraturi. Acestea sunt mai ordonate decat moleculele individuale. De ce totusi nu este contrazis principiul al doilea al termodinamicii?
3. Procesul de formare a unei micele necesita energie? De ce?
4. Explicati asemanarile si deosebirile dintre un canal ionic si un ionofor formator de pori.
5. Ce s-ar intampla daca nu ar exista transport membranar pasiv? Dar transport activ?
Alegeti raspunsul corect:
1. In sangele unui turist care a fost muscat de un sarpe au fost detectate trei enzime:
- fosfolipaza, care degradeaza fosfolipidele;
- neuraminidaza, care inlatura carbohidratii de la suprafata membranei plasmatice;
- proteaza, care degradeaza proteinele.
Care dintre aceste enzime credeti ca este responsabila pentru hemoliza eritrocitelor din sangele turistului si de ce?
a. Neuraminidaza distruge glicocalixul bogat in carbohidrati, determinand ruperea membranei eritrocitare, deoarece acesta are rolul principal in a conferi rezistenta structurala membranei.
b. Proteaza degradeaza proteinele membranare, conducand la pierderea integritatii membranei prin rupere.
c. Fosfolipaza degradeaza fosfolipidele, componenta membranara principala care stabileste bariera fata de mediul extracelular.
2. Intr-un mediu apos, lipidele formeaza o structura ordonata (micela/bistrat). Ce anume duce la aparitia acestor structuri?
a. Fosfolipidele sunt foarte ordonate in apa, iar prin formarea de micele/bistraturi isi cresc libertatea de miscare.
b. In prezenta lipidelor, moleculele de apa tind sa formeze cat mai multe legaturi de hidrogen, ceea ce se realizeaza prin minimizarea suprafetei de interactie dintre apa si regiunea hidrofoba a lipidelor.
c. Fosfolipidele au o mare afinitate unele fata de altele, ceea ce determina autoasamblarea acestora in bistraturi sau micele.
3. Capacitatea unei membrane celulare de a permite transportul unor molecule hidrofile prin membrana este determinata de:
a. proteine membranare periferice;
b. proteine membranare integrale;
c. carbohidrati membranari;
d. lipide membranare.
Multi hormoni sunt transportati in sange legati de proteinele plasmatice. Hormonii proteici si polipeptici sunt in general transportati liber. Alti ho [...] |
Multi hormoni sunt transportati in sange legati de proteinele plasmatice. Hormonii proteici si polipeptici sunt in general transportati liber. Alti ho [...] |
Dupa ce hormonii tiroidieni sunt secretati in sange, ei se leaga reversibil de proteine transportoare serice. Cantitatea de hormonii legati sau [...] |
Copyright © 2010 - 2025
: eSanatos.com - Reproducerea, chiar si partiala, a materialelor de pe acest site este interzisa!
Informatiile medicale au scop informativ si educational. Ele nu pot inlocui consultul medicului si nici diagnosticul stabilit in urma investigatiilor si analizelor medicale la un medic specialist.
Termeni si conditii - Confidentialitatea datelor - Contact