FACTORII DE REZISTENTA A ORGANISMULUI INFECTAT
Organismul este expus in toate etapele vietii la diferite infectii, cea mai mare receptivitate intalnindu-se la copii sub 1 an si batrani. La aceste varste extreme infectiile evolueaza de obicei mai sever, datorita posibilitatilor mai reduse de aparare ale organismului. Unele boli endocrine (diabet, hipotiroidie, insuficienta suprarenala), subnutritia, carenta de vitamine, oboseala, sarcina, etc., scad rezistenta generala a organismului, favorizand aparitia diferitelor infectii.
Organismul gazda poate contribui la desfasurarea procesului infectios prin mecanisme de rezistenta nespecifica si specifica (tabel 2).
Rezistenta nespecifica, naturala, innascuta fata de agentii infectiosi este comuna tuturor indivizilor unei specii;
Rezistenta specifica, dobindita (imunitatea antiinfectioasa) se dezvolta pe parcursul vietii, ca urmare a contaminarii continue cu diversi agenti infectiosi.
|
Tipuri de rezistenta |
Exemple |
|
Naturala, inascuta, nespecifica Dobindita natural Dobindita artificial |
- Barierele externe (mecanica, chimica, biologica); - Factori interni umorali (sistemul complementului) si celulari (inflamatia, fagocitoza). - Transfer placentar de anticorpi de la mama la fat (pasiva); - Dupa contactul cu un agent infectios. - Administrare de antitoxine (pasiva); - Vaccinare (activa). |
Tabel 2: Tipuri de rezistenta (dupa Moldovan Roxana, 1997)
Mecanismele rezistentei naturale sunt distincte de cele ale rezistentei dobindite, dar, ele nu pot fi separate, deoarece se intrica si actioneaza sinergic.
1. REZISTENTA NESPECIFICA (NATURALA, INNASCUTA, DE SPECIE)
Reprezinta starea de rezistenta cu care sunt dotati toti indivizii unei specii fata de anumiti agenti infectiosi. Raspunsul de aparare nespecifica este de o intensitate relativ egala, indiferent de natura agentului agresiv bacterian, si indiferent de numarul de contacte anterioare pe care organismul le-a avut cu agentul respectiv.
Fiecare specie este genetic rezistenta la anumite infectii, dar si susceptibila la altele. Astfel, omul este singura specie care face in mod natural sifilis, scarlatina, gonoreea, animalele fiind rezistente in mod natural. Aceasta rezistenta antiinfectioasa se explica prin conditiile impropri pe care un organism le ofera microorganismului fata de care este rezistent si prin lipsa receptorilor celulari specifici pentru agentul infectios respectiv.
Dar, rezistenta antiinfectioasa prezinta variatii de rasa si variatii individuale, chiar in cadrul aceleiasi specii. Este cunoscut faptul ca in timpul unor epidemii, unii indivizi fac forme grave de boala infectioasa, altii forme fruste, iar unii chiar infectii clinic inaparente. Aceasta variatie individuala a rezistentei la infectii este determinata de structura generala a mecanismelor de aparare si in special de prezenta diferitelor antigene de histocompatibilitate.
Factorii de aparare nespecifica pot fi impartiti in: barierele cutanate si mucoase, reactia febrila, factorii interni umorali si celulari.
1.1. BARIERELE CUTANATE SI MUCOASE
Tegumentele si mucoasele constituie bariere mecanice, chimice si biologice, eficiente in prevenirea patrunderii microbilor in organism (tabelul 3).
|
Sediu |
Mecanisme de protectie |
|
Piele |
- Bariera mecanica - Secretia de acizi grasi - Acidul lactic, rezultat al metabolismului local - Uscaciunea relativa a pielii |
|
Cavitatea bucala |
- Enzimele (lactoperoxidaza, lizozimul si anticorpii din saliva - Actiunea de spalare a salivei |
|
Aparat respirator |
- Tusea si stranutul - Miscarea firelor de par din vestibulul nazal care capteaza microbii - Enzimele si capacitatea surfactanta a mucusului - Miscarile cililor vibratili - Macrofagele alveolare |
|
Tract digestiv |
- PH-ul gastric (1-2) - Enzimele si anticorpii din mucus - Miscarile peristaltice - Flora intestinului gros |
|
Ochi |
- Lizozimul din lacrimi - Actiunea de spalare a lacrimilor |
|
Ureche |
- Actiunea antimicrobiana a cerumenului |
|
Aparatul genitourinar |
- Ph-ul vaginal, flora locala - PH-ul urinii - Actiunea de spalare a urinii |
Tabel 3: Bariere cutaneo-mucoase (dupa Moldovan Roxana, Cosnita Monica, 1997)
a. Bariere mecanice
Sunt reprezentate de integritatea pielii si mucoaselor, integritate care este cel mai important obstacol in calea patrunderii bacteriilor dincolo de poarta de intrare. Aceasta integritate este asigurata de: straturile cornoase ale epidermului, capcanele anatomice la nivelul mucoaselor (meatele nazale), inglobarea bacteriilor in masa de mucus cu retinerea lor, miscarea cililor epiteliului cailor respiratorii superioare, indepartarea mecanica a bacteriilor prin secretiile naturale de la nivelul mucoaselor (saliva, sputa, secretii nazo-faringiene), peristaltismul intestinal.
b. Bariere chimice
pH scazut de la suprafata tegumentelor (pH=5-6), asigurat de acizii grasi cu molecula lunga din secretia glandelor sebacee, are un puternic efect nociv pentru germenii patogeni (streptococii de grup A, bacilii difterici);
lizozimul, existent in saliva, secretia lacrimala, secretia nazo-faringiana, mucusul cervical, lichidul prostatic, este o proteina cu greutate moleculara mica, cu efect de liza a bacteriilor prin desfacerea componentelor glucidice din peretele bacterian;
acidul clorhidric din sucul gastric (pH=2) are puternic efect bactericid asupra tuturor germenilor ce patrund odata cu alimentele, asigurand sterilitatea stomacului si duodenului;
secretia biliara cu efect puternic bactericid, ceea ce explica lipsa bacteriilor in duoden si prima portiune a jejunului; bila interfereaza functiile vitale ale membranei celulare avand si o actiune neutralizanta asupra unor toxine bacteriene;
pH acid al vaginului rezultat in urma metabolizarii glicogenului de catre bacilii lactici, reprezinta de asemenea un mijloc eficient de aparare contra microbilor.
c. Bariere biologice
Sunt reprezentate de flora comensala de pe tegumente si mucoase. Coexistenta in acelasi ecosistem a diferitelor specii, tulpini si variante bacteriene, ca si asocierea altor agenti (fungi, paraziti, virusuri) este marcata puternic de relatia de antagonism bacterian, care asigura o protectie biologica eficienta, cu precadere in cazul cailor respiratorii superioare, intestinului, tegumentului, vaginului (Exemplu: speciile de Lactobacillus din vagin, mentin in mod normal un pH acid, nefavorabil dezvoltarii altor bacterii).
Mecanismele prin care flora normala se opune multiplicarii florei patogene sunt:
competitia pentru acelasi receptor celular
competitia pentru un substrat nutritiv
secretia unor produsi secundari toxici
stimularea sistemului imun, cu producerea anticorpilor naturali, care vor reactiona incrucisat cu antigenele florei patogene.
Administrarea abuziva de antibiotice cu spectru larg poate distruge echilibrul dintre speciile florei normale si unele specii conditionat patogene sau patogene, cu aparitia diareei prin dezvoltarea necontrolata a unei singure specii. Exemplu: S. aureus, Candida albicans, Clostridium difficile, care fac parte din flora normala si care se mentin la individul sanatos in anumite limite tocmai datorita antagonismului bacterian.
1. REACTIA FEBRILA
Ea insoteste, de regula, o infectie bacteriana. Implica o dereglare a mecanismelor sistemului termoreglator (central si periferic), cu predominanta termogenezei. Centrii termoreglatori din hipotalamus vor fi excitati de catre diferiti factori pirogeni ca:
endotoxinele bacteriilor Gram negative (lipopolizaharidul din peretele celular al acestor germeni actioneaza asupra macrofagelor ce vor secreta pirogeni endogeni: interleukina-l (IL-l), factorul de necroza tumoral (TNF-alfa);
alti pirogeni endogeni secretati de macrofage, monocite, granulocite, ca urmare a actiunii altor bacterii, virusuri, hormoni steroizi, limfocite T, complexe imune circulante (ura 3).
Reactia febrila, din punct de vedere al apararii antiinfectioase nespecifice, asigura mobilizarea mijloacelor de lupta ale organismului, prin activarea metabolismului si a circulatiei sanguine. O temperatura de 38,5 C accentueaza distrugerea agentilor infectiosi, prin cresterea productiei de imunoglobuline si a activarii fagocitozei. Insa, febra prelungita poate fi asociata cu efecte nevaforabile (cresterea activitatii cardiace, la un pacient cu functie cardiovasculara compromisa, poate determina aparitia convulsiilor; modificarile metabolice pot duce la deshidratare si pierdere de electroliti; cresterea susceptibilitatii la efectele unor toxine microbiene).

ura 3: Reactia febrila (dupa C. Voiculescu, 1996)
1.3. FACTORII INTERNI AI APARARII NESPECIFICE
In cazul in care bacteriile patogene au invins barierele mucoase sau cutanate, difuzarea lor este impiedicata in mod normal, de tesutul conjunctiv intercelular. Uneori insa, acesta poate fi distrus de unele enzime bacteriene proteolitice: proteinaze, fibrinolizine, colagenaze, streptochinaze. Factorii interni ai apararii nespecifice sunt incadrati in doua categorii: factori umorali si factori celulari.
a. FACTORI UMORALI
a.1. Polipeptidele bazice din unele tesuturi au efect bactericid (patrund prin peretele bacterian si altereaza functiile osmotice a membranei citoplasmatice, ducand in final la moarte celulara). Exemple: polipeptidele bazice cu efect asupra bacilului carbunos, spermina si spermidina cu efecte asupra bacilului Koch si stafilococului aureu.
a. Lizozimul este o mucopeptidaza care hidrolizeaza legaturile peptidoglicanului din peretele celular al bacteriilor, acestea devenind sensibile la liza osmotica. Actioneaza puternic asupra bacteriilor Gram pozitive. Bacteriile Gram negative nu sunt sensibile la actiunea lizozimului, deoarece peptidoglicanul este acoperit de membrana externa. Daca aceste bacterii sunt supuse initial actiunii complementului, acesta va leza membrana externa dezvelind peptidoglicanul, sensibil acum la actiunea lizozimului.
a.3. Splenocitina din splina care actioneaza pe bacilii Gram negativi: Salmonella, E. coli patogen.
a.4. Betalizina, proteina bazica de origine trombocitara, cu efect inhibitor asupra multiplicarii florei Gram pozitive, cu efect distructiv la nivelul membranei celulare.
a.5. Lactoferina este o proteina care leaga fierul si care este in competitie cu bacteriile carora le este esential in multiplicare.
a.6. Opsoninele sunt substante care adera de suprafata unui microorganism facandu-l accesibil fagocitozei. Exista opsonine specifice (anticorpi de tip IgG implicati in apararea antiinfectioasa dobandita) si nespecifice (C3b, fibronectina). Exemplu: C3b, produsa in timpul activarii complementului, se leaga de glicoprotein-receptorul pentru C3b prezent pe membrana fagocitelor. Fiind legat de fagocit microbul va fi fagocitat eficient.
a.7. Stress-ul infectios antibacterian ce consta din punerea in activitate a axului hipotalamus hipofiza anterioara corticosuprarenala cu eliberare de glucocorticoizi. Insa, in cantitate mare, glucocorticoizii scad mijloacele de aparare ale organismului (ura 4).

ura 4: Stress-ul antiinfectios (dupa C. Voiculescu, 1996)
a.8. Proteina C reactiva isi trage numele de la faptul ca reactioneaza, printre altele, cu proteina C a pneumococului. Aceasta proteina se ataseaza pe unele bacterii sau fungi si promoveaza invelirea acestora cu molecule de complement (C Invelirea celulelor cu fractiuni de C produce facilitarea fagocitarii celulelor respective.
a.9. Fibronectina, glicoproteina care favorizeaza adeziunea celulelor de suprafete, actioneaza ca opsonina fata de bacteriile Gram pozitive. Ea se gaseste la suprafata mucoaselor, impiedicand, prin proprietatea de a se lega de flora Gram pozitiva, colonizarea mucoaselor cu flora Gram negativa (de exemplu mucoasa faringiana).
a.10. Sistemul complementului (C , alexina)
Complementul este o componenta complexa a serului normal. Este format din aproximativ 25 de proteine plasmatice (componente) care se activeaza succesiv una pe cealalta intr-o ordine data (activare in cascada). Aceste componente sunt produse in marea lor majoritate de catre ficat. Macrofagele reprezinta al doilea sediu important de sinteza a componentelor complementului. A mai fost demonstrata sinteza limitata a unora din factorii acestuia in fibroblaste si celulele intestinale. Se comporta ca proteine de faza acuta: rata sintezei lor se accelereaza in cursul infectiilor sau altor injurii.
Cantitatea de complement prezenta in plasma este relativ constanta.
- Cresterea activitatii globale a complementului se poate produce in infectii sau procese neoplazice.
- Scaderea nivelurilor plasmatice ale complementului se datoreaza fie unui consum exagerat al acestuia (in bolile de complexe imune), fie unei sinteze reduse (cum este cazul insuficientei hepatice).
ACTIUNE BIOLOGICA
Exista 2 cai de activare a complementului, cu punct de plecare diferit, dar, care converg spre aceiasi produsi finali:
- calea clasica (comuna), a carei activare presupune prezenta anticorpilor specifici, deci un contact prealabil cu agentul etiologic respectiv. Activarea pe aceasta cale se produce, de regula, in prezenta complexelor antigen-anticorp. Astfel, bacteriile cu care organismul a mai venit in contact, patrund in organism si intalnesc anticorpii corespunzatori cu care vor forma complexe antigen-anticorp.
cate o molecula de C1q si de C1s si doua molecule de C1r formeaza unitatea de recunoastere (C1qrs) ce se va lega de complexul antigen-anticorp;
unitatea de recunoastere va cliva C4 si C2 in C4a, C4b si C2a, C2b; o molecula C1qrs poate sectiona un numar mare de molecule C4 si C2 inainte de a fi inactivata prin interventia unei molecule de reglare, numita inhibitorul lui C1 (C1 INH), care disociaza C1qrs;
componentele C4b si C2a vor forma C4b2a ce reprezinta C3 convertaza caii clasice, care va cliva C3 in C3a si C3b. C3b este piesa esentiala in reactiile urmatoare (ura 5).
- calea alternativa, mai veche din punct de vedere filogenetic, se activeaza in absenta anticorpilor specifici, deci face parte din rezistenta antiinfectioasa naturala. Activarea pe aceasta cale este declansata de mai multi factori: polizaharide, acizi theicoici, bacterii Gram pozitive sau Gram negative, endotoxine bacteriene, paraziti (tripanozoma), levuri (Candida albicans), agregatele de imunoglobuline (agregate termic, nu prin complexare cu antigene).
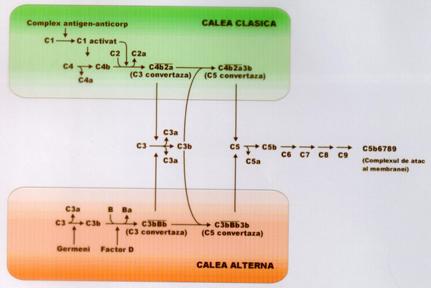
ura 5: Sistemul complementului
In plasma normala C3 este supus permanent unei activari spontane discrete prin clivarea in C3a si C3b.
C3b se combina cu factorul B (enzima proteolitica), rezultand C3bB;
dupa legarea sa de substrat, factorul B este clivat proteolitic in fragmentele Ba si Bb de o enzima prezenta in plasma in stare activa, factorul D; in urma actiunii factorului D se va obtine Ba iar C3bB va deveni C3b2Bb;
C3b2Bb este C3 convertaza caii alternative. Aceasta convertaza, rezultata in mod fiziologic, ar putea cliva cantitati mari de C3, daca nu
FORMAREA COMPLEXULUI DE ATAC AL MEMBRANEI
Deci, ambele cai produc enzime specifice (C3 convertaze) care vor actiona asupra componentei C3 a sistemului (elementul crucial al cascadei), cu scindarea sa in doua fragmente: C3a si C3b.
C3a, agent inductor de inflamatie, are proprietati chemotactice si anafilatoxice (degranularea mastocitelor cu eliberarea histaminei si cresterea permeabilitatii vasculare).
C3b, care se depune pe bacterii sau pe alte celule tinta, poate produce eliminarea acestora prin doua mecanisme: imunoaderenta si liza. O serie de fagocite (granulocitele neutrofile, macrofage) exprima pe suprafata receptori specifici pentru C3b. C3b mijloceste pe aceasta cale atasarea celulelor tinta pe fagocite (imunoaderenta) si ingurcitarea lor ulterioara. Aceasta invelire cu C a tintelor, care pregateste si faciliteaza fagocitoza, a fost denumita opsonizare.
In acelasi timp, C3b, poate media continuarea activarii in cascada a sistemului, care se va finaliza prin asamblarea unui complex enzimatic (C5, 6, 7, 8, 9) pe suprafata celulei tinta. Acest complex a fost denumit complex de atac a membranei deoarece provoaca solutii de continuitate care duc la liza celulei atacate (bacterie, celula infestata de virus sau alte organisme invadatoare).
Etapele principale ale formarii complexului de atac al membranei sunt:
ambele C3 convertaze se combina cu C3b formand C5 convertaza (C3b2BbP pentru calea alternativa si C3b4b2a pentru calea clasica);
C5 convertaza sectioneaza C5 cu generare de C5a (anafilatoxina) si fragmente active C5b;
C5b se fixeaza pe suprafata celulei (bacteriei). De remarcat este faptul ca moleculele C5b sunt foarte instabile si sunt inactivate rapid daca raman libere in plasma; in schimb, ele devin stabile daca reusesc sa se ataseze unui substrat sau membrana celulara, pe care incepe procesul de asamblare a complexului de atac;
de C5b se vor lega in continuare C6, C7, C8 care se gasesc libere in ser si se asambleaza pe molecula C5 numai in momentul activarii ei, fara interventia vreunei enzime proteolitice. Se formeaza complexul C5b678, care are capacitate litica redusa. Celulele tinta pot fi lizate doar atunci cand densitatea unor asemenea complexe este foarte mare;
in continuare, mai multe molecule de C9, care polimerizeaza, se ataseaza de complexul C5b678, ducand la cresterea considerabila a puterii litice. Structura proteinei C9 este asemanatoare cu a perforinelor prezente in granulatiile celulelor citotoxice;
se formeaza, astfel, un complex de atac al membranei celulare (C5b6789) cu liza consecutiva a celulei bacteriene.
Sunt reprezentati de fagocitoza si inflamatie.
b.1. FAGOCITOZA
Constituie un proces esential, prezent pe toata durata infectiei. Reprezinta reactia de aparare celulara a organismului si consta dintr-un mecanism de indepartare a microorganismelor si a celulelor lezate, prin inglobarea lor de catre anumite celule ale organismului (fagocite) specializate pentru aceasta functie.
Aceste celule migreaza la locul de patrundere al bacteriilor in organism, le inglobeaza si le distrug in interiorul lor, prin fagolizozomi (enzimatic). Se realizeaza in acest fel localizarea infectiei, care clinic se manifesta prin inflamatie cu simptomele caracteristice: rubor, tumor, calor, dolor, si functio laesa. In acest fel o infectie minora poate fi invinsa, printr-o buna aparare locala, iar puroiul rezultat din resturile celulelor distruse, contine bacterii moarte (puroi steril). De cele mai multe ori insa, infectia progreseaza, determinand diferite manifestari clinice.
CATEGORII DE CELULE CU PROPRIETATI FAGOCITARE (FAGOCITE)
In fagocitoza intervin doua categorii principale de celule: microfagele si macrofagele. Aceste celule au capacitatea de a migra dirijat spre locul unde sunt particule straine. Migrarea lor in tesuturi este favorizata de substantele chemotactice: C5a, C5b si alti factori produsi in cursul inflamatiei. Ele au pe suprafata lor receptori pentru fragmentul Fc al imunoglobulinei G (IgG) si receptori pentru fractiunea C3b a complementului si pot dezvolta mecanisme puternic microbicide. Se mai numesc si fagocite profesioniste, pentru a putea fi deosebite de celulele fibroblaste, endoteliale si reticulare care alcatuiesc fagocitele neprofesioniste; acestea indeplinesc o activitate fagocitara mai redusa, deoarece sunt lipsite de receptorii prezenti pe fagocitele profesioniste si mai ales nu dezvolta mecanisme bactericide.
- Granulocitele neutrofile realizeaza fagocitarea corpilor bacterieni integri sau fragmentelor de germeni. Capacitatea de fagocitoza este marita de procesul de opsonizare, opsoninele fiind anticorpi de tip IgG. Dupa inglobare, antigenele straine sunt distruse datorita: enzimelor lizozomale si lactoferinei (din granulatiile specifice neutrofile); mieloperoxidazei, elastazei, catepsinei, beta-glucuronidazei (din granulatiile azurofile). Ele sunt celule kamikaze ale apararii antiinfectioase, deoarece sosesc primele la locul injuriei. Dupa digestia materialului fagocitat, foarte frecvent, leucocitele sunt distruse.
- Polimorfonuclearele eozinofile iau parte la apararea antiparazitara. Contin enzime ce metabolizeaza histamina si leucotrienele fiind astfel capabile sa joace rol important in limitarea reactiilor alergice. Ele contin si proteine toxice (proteina majora bazica) ce duce la distrugerea celulelor epiteliale din tractul respirator ducand la inflamatie cronica in astmul bronsic. Ele fagociteaza complexe antigen-anticorp precum si unele resturi antigenice ce rezulta din raspunsul imun si care in alte conditii ar duce la boli autoimune.
- Polimorfonuclearele bazofile (echivalentul mastocitelor tisulare) au granulatii mari ce contin numerosi mediatori chimici si precursorii acestora, ca, de exemplu: histamina, prostaglandine, leucotriene, factori activatori ai trombocitelor. Acesti mediatori sunt eliberati, la nevoie, in cantitati reduse. Elaborarea lor masiva poate fi daunatoare cu aparitia reactiilor alergice de tip I: astm bronsic, urticarie, febra de fan etc.
MACROFAGELE
Pot fi mobile, prezente in sange (monocitele) si fixe, dar usor mobilizabile, ce fac parte din sistemul reticulo-endotelial (sistem fagocitar mononuclear). Exemple: macrofagele din splina si ganglionii limfatici; celulele Kuppffer din ficat; histiocitele din tesutul conjunctiv; unele celule ale nevrogliei.
In tabelul 5 sunt prezentate ativ principalele proprietati si functii ale polimorfonuclearelor si celulelor sistemului reticulo-endotelial.
|
Polimorfonucleare |
Sistem fagocitar mononuclear |
||
|
Proprietati |
Functii |
Proprietati |
Functii |
|
- Au origine medulara - Sunt celule cu viata scurta - Au pe suprafata receptori pentru C3 si Fc a IgG - Au un numar mare de lizozomi - Mieloperoxidaza prezenta in lizozomi - Sunt active neutrofilele, apoi eozinofilile |
- Sunt foarte mobile - Sunt primele care se deplaseaza la locul infectiei - Reprezinta prima linie de celule care fagociteaza - Digera total bacteriile fagocitate - Eozinofilele inglobeaza complexe antigen-anticorp |
- Origine medulara - Sunt celule cu viata lunga - Au pe suprafata receptori pentru C3 si Fc a IgG - Au un numar mic de lizozomi (numarul creste in macrofagul activat) - Nu au mieloperoxidaza - Cuprinde monocitul sanguin si macrofagele tisulare. Macrofagele difera morfologic intre ele, dar functioneaza identic |
- Fagocitoza - Functie secretorie - Celule asociate raspunsului imun |
Tabel 5: Caractere ale fagocitelor polimorfonucleare si celulelor sistemului fagocitar mononuclear (dupa A. Ivanof si M. Ciupe, 1982)
Macrofagele fagociteaza macromolecule inerte, microorganis-mele opsonizate, pe care le distrug in mare parte (tabel 6).
Unele microorganisme (micobacterii, listerii, brucele, toxoplasme) supravietuiesc si se inmultesc in macrofage. In acest caz, macrofagele protejeaza microorganismele respective, servind ca factor de diseminare a infectiei.
|
Functie |
Exprimare |
|
1. Fagocitoza Functie secretorie 3. Cooperare cu sistemul imun |
- Reprezinta a doua linie de celule care fagociteaza ; - Nu digera complet microorganismele; - Inglobeaza resturi celulare si tisulare, curatind teritoriul lezat. - Enzime cu actiune asupra proteinelor prezente in mediul extracelular: colagenaze, elastaze, proteaze lizozomale, activatori ai plasminogenului; - Secretie de factori antiinfectiosi: lizozim, fractiuni ale complementului (C2, C4, probabil C3, C5), interferon; - Factori ce modeleaza functia altor celule: proteina mitogena asupra limfocitului T si a timocitului; - Citotoxine asupra celulelor straine si tumorale; - Reglatori ai activitatii celulare: prostaglandine; - Factori de diferentiere: factorul de stimulare al coloniilor, care stimuleaza diferentierea celulei susa medulare in linia granulocitara. - Intervine in declansarea si reglarea raspunsului imun. |
Tabel 6: Functiile celulelor sistemului fagocitar mononuclear (dupa A. Ivanof si M. Ciupe, 1982)
ETAPELE FAGOCITOZEI
Sunt reprezentate de: chemotaxie, adsorbtia particulelor pe fagocit, ingestia particulelor in fagocit si digestia intracelulara.
CHEMOTAXIA
Reprezinta mobilizarea unidirectionala, dirijata a polimorfonuclearelor spre locul unde se elibereaza factori chemotactici. Acesti factori pot fi atat de origine bacteriana (formilmetionil-leucil-fenilalanina), cat si factori ai organismului gazda, ce apar in cursul inflamatiei: C5a, kalicreina produsa de tesuturile lezate, prostaglandine (E2, D2), tromboxan, leucotriene (B4), amine vasoactive, fragmente de colagen, de fibrina (tabel 7). Sub influenta factorilor chemotactici fagocitele parasesc capilarele prin diapedeza.
Pentru fiecare dintre factorii chemotactici, fagocitele au receptori specifici. In urma interactiunii dintre acesti factori si receptorii celulari prezenti pe suprafata PMN, se produc modificari ale fagocitului.
|
Componente bacteriene |
Componente ale organismului |
|
- Proteine bacteriene, indeosebi N-formil-metionil-peptide; - Unele lipide bacteriene. |
- Componente din sistemul complement: C5a, C5,6,7, obtinute prin activare; - Proteine serice: Kallikreina, fragmente de fibrina; - Histamina; - Derivati arahidonici; - Produse ale limfocitelor (limfokine); - Unii factori eliberati de neutrofile. |
Tabel 7: Factori chemotactici (dupa A. Ivanof si M. Ciupe, 1982)
ADSORBTIA (ATASAREA) PARTICULELOR PE SUPRAFATA FAGOCITELOR
Atasarea pe suprafata fagocitului poate fi intamplatoare sau dirijata. Contactul intamplator depinde de sansa de coliziune intre fagocite si particule. In cel de-al doilea caz, fagocitele migreaza catre bacterii sau alte particule datorita chemotaxiei. S-a demonstrat ca unele extracte bacteriene sau corpi bacterieni integri atrag leucocitele (chemotaxie pozitiva).
Dar, in mod normal, pentru ca sa permita atasarea fagocitului, suprafata celulei bacteriene trebuie modificata, si aceasta pregatire in vederea fagocitarii se numeste opsonizare. Opsonizarea se poate realiza prin mecanisme specifice, deoarece presupun participarea anticorpilor rezultati in urma unui raspuns imunitar. Este vorba de interventia IgG, care se leaga de fagocite prin fragmentul Fc, realizand o punte intre antigenele de suprafata si receptorul de pe fagocit, sau prin actiunea combinata a complementului si a anticorpilor, asa cum se intampla in cazul IgM. IgM nu se pot fixa pe suprafata fagocitului, dar intervin formand complexe imune care activeaza complementul, activare din care rezulta fractiunea C3b pentru care exista receptori pe fagocit (ura 6).
Opsonizarea se poate realiza si prin mecanisme nespecifice, prin interventia proteinei C reactive, a fibronectinei, C3b, alfa 2-glicoproteinei serice.
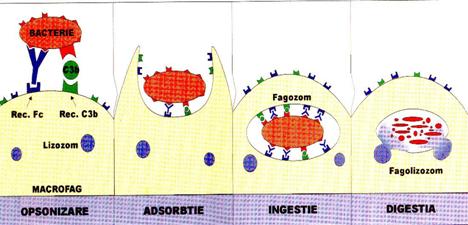
ura 6: Etapele fagocitozei
Un factor potentator al atasarii particulelor este si asa numita fagocitoza de suprafata, prin care microorganismele sunt captate mai usor daca sunt prinse pe suprafete rugoase. Aceasta se poate realiza prin plasarea microorganismelor pe suprafetele dintre leucocite, intre leucocite si suprafetele tisulare, sau in interstitiile cheagurilor de fibrina.
Adsorbtia particulelor pe suprafata fagocitului este inhibata de anumiti factori: hiperosmolaritatea mediului, concentratii crescute de glucoza si, respectiv, scazute de fosfati in mediu etc.
Dupa atasare, particulele sunt inglobate intr-o vacuola formata de membrana citoplasmica prin emitere de pseudopode. Procesul necesita energie, care provine de obicei din glicoliza. Dupa inglobarea particulei, cula fagocitara se deplaseaza spre interiorul celulei.
Etapa de ingestie (inglobare) se desfasoara cu ritmuri si intensitati diferite, in functie de interventia unor factori potentatori sau inhibitori. Din prima categorie, amintim existenta in membrana fagocitului a unor proteine contractile din familia actino-miozinei, care asigura emiterea de pseudopode si formarea pungilor. Factorii inhibitori sunt mai putin bine studiati, un rol probabil in acest sens avandu-l prostaglandinele din seria E si leucotrienele din grupul B.
DIGESTIA
Vacuola formata (fagozom) va fuziona cu lizozomii (grupati in jurul vacuolei) rezultand fagolizozomii. Formarea fagolizozomilor este un proces insotit de disparitia granulatiilor intracitoplasmatice (degranulare). Liza granulelor ar avea loc la contactul cu peretele pungii fagocitare si consta in eliberarea enzimelor lizozomale, care sunt diverse substante cu actiune microbicida, care vor declansa digestia. In acelasi timp, are loc o activare puternica a metabolismului oxidativ al PMN.
Distrugerea microbilor se petrece prin 2 mecanisme: O2-dependente si O2-independente.
Mecanismele O2-dependente
Sunt consecinta exploziei respiratorii, o cale metabolica de respiratie in stare dormanta intr-o celula inactiva. Intensificarea brusca a metabolismului (explozia respiratorie) duce la formarea unor compusi de tipul: ionilor de superoxid (O3-), oxigen atomic (O-), radicali hidroxili, peroxid de hidrogen (H2O2) si hipoclorit (HOCl). Toti acesti produsi au efect puternic bactericid.
Fenomenul digestiei intracelulare a particulelor are o complexitate deosebita. In esenta, au loc urmatoarele secvente:
eliberarea enzimelor lizozomale din fagolizozomi in interiorul vacuolei;
eliberarea in vacuola si a altor enzime celulare: mieloperoxidaza, o enzima foarte activa stocata in PMN, care in mediu acid existent in fagolizozom duce la formarea unor halogeni cu efect puternic microbicid;
activarea proceselor oxidative si anume: utilizarea intensa de catre celula a oxigenului molecular, care, sub actiunea catalitica a NADPH (nicotinadenindinucleotidfosfat redus), se transforma in superoxid; superoxidul este clivat, datorita superoxid-dismutazei, in oxigen molecular si peroxid de hidrogen;
peroxidul de hidrogen, in cooperare cu mieloperoxidaza, determina dezagregarea particulelor fagocitate; peroxidul de hidrogen eliberat in fagocit este un agent bactericid puternic, dar combinarea cu mieloperoxidaza ii creste de 50 de ori activitatea.
Mecanismele bactericide independente de O2
Aceste mecanisme care intervin in omorarea microorganismelor, se datoreaza unor enzime:
enzime hidrolitice (catepsina, glicozidaza, arilsulfataza) care digera peretele celular bacterian;
defensine: proteine cationice care se leaga de peretele celular si determina formarea unor canale ce strapung peretele, atat la bacteriile Gram pozitive, cat si la cele Gram negative;
lizozimul, prezent in granulatiile lizozomale, ce ataca peptidoglicanul;
histone nucleare sau pH-ul acid din fagolizozom, care impiedica viata microorganismului fagocitat;
lactoferina care spoliaza mediul de fier necesar bacteriilor pentru multiplicare.
In digestia intracelulara a particulelor mai pot interveni si mecanisme aditionale, ca: aparitia in citoplasma a ionului de clor oxidat, cu puternica actiune bactericida; folosirea de catre celula a caii scurtate hexozo-monofosfat, in vederea sintezei ulterioare de superoxid (O2), peroxid de hidrogen (H2O2), acidul hipocloros (HOCl).
Omorarea microorganismelor fagocitate este urmata de digerarea completa a acestora prin activitatea unor hidrolaze. In PMN se produce degradarea completa a microorganismului, spre deosebire de macrofage, in care particula fagocitata este degradata partial, incomplet, antigenele fiind apoi prezentate sistemului imun.
Calea hexozomonofosfatilor
![]()
NADPH Superoxid Mielo
![]()
![]()
![]() O2 O2 H2O2 HOCl
O2 O2 H2O2 HOCl
oxidaza dismutaza peroxidaza
b. INFLAMATIA
Este un proces natural de aparare antiinfectioasa, un mecanism important de rezistenta naturala, dar care este reglat in evolutie de procese imune si de foarte multe ori procesul inflamator este dependent de factori imunitari.
Consta dintr-un ansamblu de procese clinice si fiziopatologice de aparare celulara nespecifica, declansat de diferiti agenti, cu deosebire infectiosi, dar si de iritanti cum ar fi substantele chimice, caldura puternica si leziuni mecanice. Rezultatul acestui proces este acumularea unui mare numar de celule fagocitare la locul inflamatiei.
Inflamatia este un mecanism de aparare antiinfectios rapid, care tinde sa localizeze infectia si sa previna diseminarea ei. Manifestarile clinice sunt eritemul (rubor), caldura (calor), edemul (tumor), durerea (dolor) si impotenta functionala a regiunii respective (functio laesa).
Inflamatia poate evolua spre: vindecare cu restitutio ad integrum a tesutului sau vindecare cu sechele. Deznodamantul unei reactii inflamatorii depinde de extinderea procesului inflamator, de microorganismele implicate, precum si de reactivitatea gazdei.
ETAPELE INFLAMATIEI (ASPECTE FIZIOPATOLOGICE)
VASODILATATIE CAPILARA LOCALA
Inrosirea zonei inflamate este rezultatul cresterii cantitatii de sange la acest nivel. Dilatatia vaselor sanguine locale determina o scadere a circulatiei sangelui,care este insotita de o crestere a permeabilitatii capilarelor locale ceea ce cauzeaza aparitia edemului.
ATRACTIA LEUCOCITELOR (CHEMOTACTISM)
Chemotactismul este reprezentat, in acest caz, de atractia intre leucocite si spatiul interstitial. Primele celule care ajung in tesutul agresat sunt PMN, datorita capacitatii mari de migrare, apoi celulele sistemului reticuloendotelial si limfocitele care au rol principal in initierea reactiilor imunitare.
Toate aceste celule sunt inzestrate cu o mare capacitate de receptie a stimulilor chemotactici, care sunt reprezentati in mare, de factorii chemotactici de natura bacteriana, de factorii produsi in urma activarii C , si nu in ultmul rand, de cei rezultati din degranularea mastocitelor. Cei mai importanti factori care asigura producerea inflamatiei sunt: aminele vasoactive (de tip histaminic), serotonina, unele enzime proteolitice (enzime lizozomale, kalicreina), unele subfractiuni de complement (C3a, C5a) denumite anafilatoxine (ce determina degranularea mastocitelor cu eliberarea unor mediatori chimici), interleukina-l (IL-l) si interleukina-8 (IL-8) ce sunt monokine secretate de macrofag. O parte din acestia sunt preformati, asa cum este histamina, cu efect vasodilatator asupra capilarelor si cu cresterea permeabilitatii acestora.
Sub actiunea acestor factori, PMN vor parasi capilarul prin diapedeza si se vor indrepta catre focarul inflamator.
In consecinta se produce o exudare a plasmei cu mediatorii pe care aceasta ii contine, din capilare spre zona infectata (cresterea fluxului sanguin spre zona inflamata, cu cresterea temperaturii locale).
Sub actiunea unor mediatori chimici rezultati in timpul activarii C si secretati de macrofagele stimulate de toxinele bacteriene, se modifica endoteliul capilar care permite adeziunea PMN de acesta.
DIAPEDEZA LEUCOCITARA
Este procesul prin care un fagocit se strecoara printre celulele endoteliului vascular, trecand in spatiul interstitial. Aceasta trecere a celulelor in spatiul interstitial se face printr-un mecanism cu mai multe etape: rostogolire, marginatie-adeziune, traversare prin emiterea de pseudopode (ura 8).
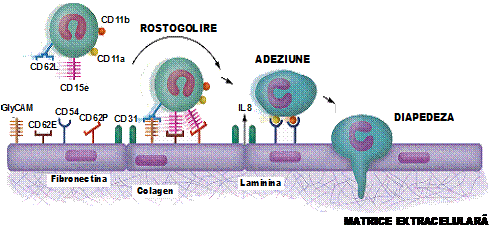
ura 8: Etapele inflamatiei
In procesul de trecere a fagocitelor prin celulele endoteliului vascular intervin moleculele de adeziune, ce fac parte din trei familii: selectine, integrine si superfamilia imunoglobulinelor. Moleculele de adeziune sunt structuri exprimate pe suprafata leucocitelor si a celulelor endoteliale care permit, prin interactiuni reciproce, atasarea leucocitelor de endoteliul capilar, cu o forta crescanda, facilitand, in final, diapedeza. Exista doua categorii de molecule de adeziune (MA): L-selectine (CD 62L, LAM-l) sunt MA exprimate pe granulocite; E-selectine (CD 62E, ELAM-l), P-selectine (CD 62P), ICAM-l (CD 54) ce sunt MA exprimate pe endoteliile vaselor mici.
Initial, prin L-selectine granulocitele se leaga de membrana celulelor endoteliale, legare slaba, sub actiunea curentului sanguin granulocitele ruleaza (se rostogolesc) pe suprafata endoteliului vascular. Sub actiunea unor atractanti (IL-8) se modifica forma celulelor atasate, ceea ce determina oprirea rularii si adeziunea ferma la endoteliu. Aceeasi IL-8 de pe suprafata endoteliului distruge L-selectinele, cu modificarea formei celulelor facandu-le apte sa traverseze peretele vascular.
PMN vor parasi capilarul prin diapedeza si se vor indrepta catre focarul inflamator unde vor distruge microbii prin fagocitaza. Formarea puroiului poate fi asociata inflamatiei. Dupa ce fagocitele au distrus celulele bacteriene, sunt degradate si mor. In aria afectata se formeaza o masa de celule distruse, fagocite moarte, ceea ce reprezinta puroiul.
CONCLUZII
In esenta, organismul este aparat fata de unele infectii prin zestrea sa ereditara, care nu permite dezvoltarea anumitor germeni. Germenilor la care este sensibil, organismul se opune patrunderii lor prin bariere externe anatomice si chimice la care se adauga flora normala a organismului (antagonism bacterian). Daca totusi germenii au reusit sa strabata aceste bariere si ajung in zonele interne sterile ale organismului, intervin o serie de factori umorali, dintre care unul este de o importata majora: complementul. Activarea complementului are ca urmare liza celulelor microbiene, dar si declansarea raspunsului inflamator infectios. Cascada de activare a complementului va antrena eliberarea unor mediatori chimici care participa direct in procesul inflamator si vor atrage in focarul infectios un aflux mare de celule fagocitare (PMN, macrofage). Acestea vor distruge microorganismele prin fagocitoza.
Insa, in acelasi timp, exista germeni care dezvolta strategii ce ocolesc mecanismele rezistentei naturale si care pot da infectii la indivizii neimunizati. Astfel de strategii se refera la inhibitia fagocitozei (secretia de toxine fata de fagocite; rezistenta fata de lizozim; inhibitia exploziei respiratorii a fagocitelor) sau la rezistenta la actiunea complementului (inactivarea C5a la Pseudomonas aeruginosa; prezenta la suprafata bacteriilor a unor substante - acidul sialic - care determina degradarea C3b la E. coli; impiedicarea insertiei complexului de atac al membranei la Salmonella, E. coli).
Sintetizand datele privind factorii, respectiv mecanismele de aparare nespecifica ale organismului, putem constata ca prezenta factorilor nespecifici umorali sau celulari in diferite umori, sange, tesuturi si organe, constituie un sistem de aparare dinamica si interdependenta, prin care organismul uman raspunde la atacul microorganismelor. Acest sistem cuprinde sistemul fagocitar (polinucleare si mononucleare), sistemul kininelor declansator al proceselor de inflamatie, sistemul complementului si sistemul de coagulare si fibrinoliza.
O problema majora in Romania este poluarea atmosferei provocata de degajarea in aer a gazelor si particulelor fine (cenusii) prin furnale, turnurile s [...] |
Fumatul provoaca, favorizeaza sau inrautateste direct sau indirect o serie de afectiuni. La un consum zilnic de 20 de tigari, fumatorii au [...] |
FACTORII DE REZISTENTA A ORGANISMULUI INFECTAT Organismul este expus in toate etapele vietii la diferite infectii, cea mai mare receptivitate [...] |
Copyright © 2010 - 2026
: eSanatos.com - Reproducerea, chiar si partiala, a materialelor de pe acest site este interzisa!
Informatiile medicale au scop informativ si educational. Ele nu pot inlocui consultul medicului si nici diagnosticul stabilit in urma investigatiilor si analizelor medicale la un medic specialist.
Termeni si conditii - Confidentialitatea datelor - Contact