STRUCTURA CELULEI BACTERIENE
Unitatea morfofunctionala a bacteriilor este celula. Bacteriile au, ca tip de celula, celula procariota, care se deosebeste de celula eucariota prin structura si organizarea ei. Diferentele esentiale intre celula eucariota si celula procariota sunt prezentate in tabelul 1.
|
Caracter |
Celula eucariota |
Celula procariota |
|
Dimensiuni |
- mari (10 - 100m |
- mici (1 - 10m |
|
Organizare |
- multicelulara, de obicei, cu diferentieri functionale tesuturi |
- unicelulara (populatii) |
|
Modalitati de reproducere |
- mitoza sau meioza |
- diviziune directa (fisiune binara sau sciziparitate) |
|
Nucleu |
- cromozomi 2n (diploid) sau n (haploid) - nucleol prezent - membrana nucleara prezenta (material ADN concentrat) |
- cromozom unic, inelar - molecula ADN dublu helix - nucleol absent - membrana nucleara absenta (material ADN dispus difuz cu concentrare maxima la centru) |
|
Membrana citoplasmatica |
- contine steroli |
- nu are in structura steroli (exceptie genul Mycoplasma) |
|
Citoplasma |
- este timentata - organite tubulare prezente: reticul endoplasmatic, mitocondrii, aparat reticular intern Golgi, lizozomi - ribozomii fixati pe reticulul endoplasmatic rugos, sunt de 80 S cu subunitati de 40 S si 60 S |
- este netimentata - organite tubulare absente - ribozomi liberi in citoplasma, sunt de 70 S cu subunitati de 30 S si 50 S - organite particulare prezente: mezozomi, oxizomi, plasmide |
|
Strat periferic corp celular |
- membrana celulara |
- perete celular |
|
Elemente anexa |
- cilii (rar) |
- cili, fimbrii, capsula |
|
Forme de rezistenta |
- nu exista |
- spori |
Tabel 1: Diferente intre celula eucariota si procariota (dupa V. Bilbie si N. Pozsgi 1984)
Bacteriile au o structura foarte complexa, fiind alcatuite din: componente obligatorii si componente facultative.
Componentele obligatorii sau elementele intrinseci sunt prezente la toate speciile bacteriene si sunt reprezentate de: nucleu, citoplasma, membrana citoplasmatica si perete celular.
Componentele facultative sau elementele anexa sau extrinseci se gasesc doar la unele specii bacteriene si sunt reprezentate de cili sau flageli, fimbrii si capsula.
Desi mult mai mici si mai simple decat celulele eucariote, bacteriile sunt asemanatoare cu acestea in ceea ce priveste mecanismele functionale celulare, prezentand o organizare complexa cu o arhitectura externa si interna distincta (ura 2).
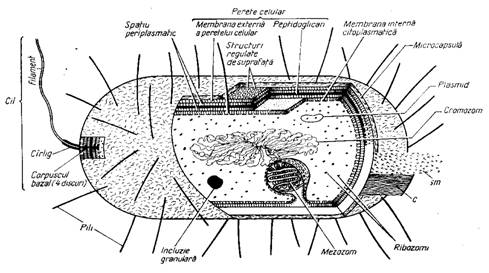
ura 2: Schema generala de organizare a celulei bacteriene (dupa V. Bilbie si N. Pozsgi 1984)
2.1. Elemente anexa (extrinseci)
2.1.1. CILII (FLAGELII) BACTERIENI
Cilii sau flagelii sunt formatiuni filamentoase ale speciilor microbiene mobile, foarte lungi, subtiri, fragile. Sunt structuri helicoidale, prezenti mai ales la bacili (familia Enterobacteriaceae) dar si la unii coci (enterococ).
Sunt formatiuni implicate in locomotie, a caror existenta este controlata genetic. Astfel, la E. Coli s-a stabilit ca pentru sinteza cililor coopereaza aproximativ 30 de gene, a caror informatie este necesara biosintezei constituentilor structurali ai cilului, in asamblarea acestora, precum si in chemotaxie si mobilitate.
Dispozitia si numarul cililor sunt caracteristice speciei (ura 3). S-au descris bacterii atriche (fara cili), monotriche - cu un cil polar (Vibrio cholerae) sau subpolar (enterococ), lofotriche - cu un smoc de cili situat la unul din polii bacteriei (Pseudomonas fluorescens), amfitriche - cu cilii situati la ambii poli ai bacteriei (la genul Spirillum) si peritriche - cu cilii dispusi pe intreaga suprafata a bacteriei (Salmonella, E. coli, Proteus, etc.).
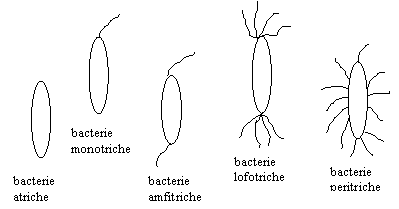
ura 3 : Tipuri de celule bacteriene in raport cu dispozitia cililor (dupa A. Ivanof si M. Ciupe, 1982)
Din punct de vedere al compozitiei chimice, cilii contin o proteina contractila denumita flagelina, asemanatoare cu miozina din celula musculara animala. In compozitia flagelinei, a fost descris un aminoacid caracteristic N-metil-lizina, care apare in cursul sintezei flagelului, prin modificarea structurii chimice a lizinei, dupa incorporarea acesteia in proteina flagelara.
Motilitatea cililor are la baza eliberarea de legaturi macroergice prin descompunerea ATP ADP + radical fosforic (acceptor de radical fosforic este molecula de arginina).
Cilii se evidentiaza la microscopul optic numai pe fond intunecat sau dupa coloratii speciale (precedate de tehnici de tratare pentru precipitarea proteinelor flagelare si mordansare). Structura intima a cililor bacterieni este furnizata de microscopia electronica. Astfel a fost descrisa existenta a trei componente morfologic distincte: corpul bazal, carligul si filamentul terminal (ura 4).
- Corpul bazal (granulatia bazala) reprezinta componenta cilului prin care acesta se ataseaza la corpul celulei bacteriene. Este o componenta structurala complexa, montata in intregime in peretele celular si membrana citoplasmatica. Este constituit din patru discuri paralele, dispuse sub forma a doua perechi pe o tija care trece prin centrul lor.
La bacteriile Gram negative exista doua perechi de discuri ce delimiteaza corpul bazal: o pereche interna (inele M si S) localizata in membrana citoplasmatica si o pereche externa situata spre carlig (inele L si P) in care discurile sunt mai distantate. Cele patru discuri au fost denumite M, S, P si L dupa presupusa lor atasare la nivelul membranei citoplasmatice, spatiului periplasmatic, stratului de peptidoglican si respectiv stratului lipopolizaharidic.
La bacteriile Gram pozitive exista o singura pereche de discuri ancorata in membrana citoplasmatica.
La nivelul corpului bazal se afla motorul ciliar, a carui miscare este transmisa proteinei flagelare, ceea ce face ca cilul sa se invarteasca in sens orar (rostogolirea bacteriei cu schimbarea directiei de miscare) sau antiorar (deplasarea bacteriei in linie dreapta).
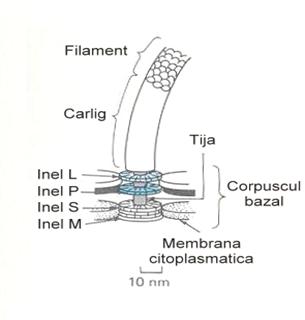
ura 4: Structura cililor (dupa George A. Wistreich, Max D. Lechtman 1988)
- carligul cilului - un manson indoit in unghi drept;
- filamentul cilului - liber in afara celulei, cu lungime de 15-25 m
Cilii sunt organe de locomotie pentru bacterii. Mobilitatea si directia miscarii cililor este asociata cu proprietatea de chemotaxie, care este o miscare dirijata inspre sau dinspre o substanta chimica. Exista o chemotaxie pozitiva care se refera la miscarea spre o concentratie pozitiva a unei substante chimice (zaharuri, aminoacizi) si aceasta se intalneste in mod obisnuit cand substanta chimica reprezinta un avantaj pentru celula (substanta nutritiva). Se vorbeste si de o chemotaxie negativa, o miscare prin care bacteria se departeaza de o substanta chimica (fenolul, acizii, bazele), de obicei cand aceasta este daunatoare, toxica pentru celula. Stimulii chimici care induc chemotaxia pozitiva sunt numiti atractanti, iar cei care induc chemotaxia negativa - repelenti.
Cilii sunt sediul antigenelor flagelare (H), importante in identificarea bacteriilor (exemplu Salmonella), stimuland un raspuns imun. In plus, pot indeplini rolul de receptori pentru virusuri si in unele cazuri determina aderarea de epitelii (la vibrionul holeric cilul intervine in aderarea de epiteliul intestinal).
In practica de rutina nu se evidentiaza flagelii, ci mobilitatea bacteriilor, fie prin examinarea lor pe un preparat nativ intre lama si lamela in care se urmaresc miscarile bacteriilor, fie prin insamantarea tulpinii pe un mediu semisolid prin inteparea in profunzime a mediului. Daca bacteria creste numai pe traseul de insamantare este imobila, daca difuzeaza in mediu, ea este mobila.
2.1.2. FIMBRIILE (PILII)
Fimbriile sau pilii sunt prelungiri scurte, rigide si groase evidentiate mai ales la speciile bacteriene Gram negative (speciile familiei Enterobacteriaceae, Neisseria gonorrhoeae) si mai putin la bacteriile Gram pozitive (Streptococcus, Corynebacterium). S-a constatat ca in cadrul aceleasi specii pot exista tulpini fimbriate si tulpini nefimbriate. Sunt elemente mai rezistente decat flagelii, in numar de 100-500/celula, cu dispozitie peritricha, cu evidentiere numai prin microscopie electronica.
Din punct de vedere chimic sunt polimeri proteici de pileina care rezista la tripsina, pepsina, acizi, baze. Aceasta natura proteica a pililor le confera proprietati antigenice.
Functional, pilii se impart in doua categorii:
- pili comuni sau pili de aderenta (fimbri) a caror sinteza este controlata de gene cromozomiale. Se gasesc in numar de 100-200 pe suprafata celulei si au rol in aderarea de diferite suprafete, in special epitelii, de unde si denumirea de adezine, care initiaza procesul infectios. Pilii comuni constituie un factor important de virulenta (de exemplu la gonococ). In afara de aderenta, acesti pili mai au si proprietati antifagocitare.
- pilii de sex (sex pilii) sunt formatiuni putin mai lungi, flexibile, cu structura si forma diferita (sferica, forma de cupa, de disc), in numar de 1-4 pe celula si intotdeauna codificati de formatiunile genetice extracromozomiale - plasmide. Acesti pili prezinta o importanta deosebita in transferul de material genetic intre bacterii, formand punti intre celula donatoare si cea receptoare, in cursul procesului de conjugare. Sunt prezenti mai ales la bacteriile Gram negative (Enterobacteriaceae, Pseudomonas).
Ambele categorii de pili prezinta antigene specifice piliare si pot fi receptori pentru bacteriofagi.
2.1.3. CAPSULA
Unele microorganisme secreta la suprafata un invelis extracelular ce inconjoara peretele celular. In functie de structura si de raporturile pe care le stabileste cu celula bacteriana, se poate vorbi de capsula propriu-zisa, microcapsula si strat mucos (glicocalix).
Din punct de vedere chimic, capsula tuturor bacteriilor de interes medical este de natura polizaharidica (Streptococcus pneumoniae, Klebsiella, Haemophilus, Bordetella), formand o retea stransa peste peretele celular (Streptococcus pneumoniae) sau avand o structura lamelara (Klebsiella). Mai rar, capsula poate fi de natura proteica (bacilul carbunos).
Rolul cel mai important al capsulei este legat de patogenitatea bacteriei (factor de virulenta): capsula impiedica fagocitarea bacteriilor care reusesc, astfel, sa scape de sub actiunea mecanismelor de aparare ale organismului. La speciile capsulate, pierderea capsulei are ca rezultat pierderea virulentei, fie direct, fie indirect prin efectul chemotactic negativ al substantelor pe care le contine (exemplu: Streptococcus pneumoniae, de tip S, capsulat, produce la soarecele alb de laborator o septicemie mortala, pe cand varianta necapsulata nu este patogena).
Capsula este o structura cu proprietati antigenice specifice care permit diferentierea unor serotipuri in cadrul speciei. Pe baza antigenului capsular se poate face tipizarea bacteriilor (la Str. pneumoniae polizaharidul capsular determina peste 80 de tipuri antigenice, iar la Haemophilus 6 tipuri antigenice).
Functiile capsulei
protejeaza bacteriile de diferiti agenti antibacterieni din mediu cum sunt: bacteriofagii, complementul, lizozimul sau alte enzime bacteriolitice;
protejeaza bacteriile de actiunea fagocitelor (factor de virulenta);
reprezinta sediul antigenelor capsulare, importante in identificarea acestor bacterii.
2.2.1. NUCLEOIDUL (NUCLEUL) BACTERIAN
Materialul nuclear bacterian (nucleoidul) are o organizare primitiva in atie cu nucleul celulelor eucariote, in sensul ca nu are membrana nucleara si nici nucleoli, aceasta fiind principala caracteristica structurala a celulelor procariote.
Din punct de vedere chimic ADN este constituit din baze azotate purinice (adenina-A si guanina-G), baze azotate pirimidinice (citozina-C si timina-T), un zahar (dezoxiriboza) si acid fosforic. O baza azotata purinica sau pirimidinica legata de o molecula de dezoxiriboza si de un radical fosforic reprezinta o unitate functionala numita nucleotid; nucleotidele, la randul lor, sunt unite intre ele prin componentele fosforice formand un lant polinucleotidic (catena). Structura dublu helicoidala a ADN (model propus de Watson si Crick in 1953) se realizeaza prin infasurarea a doua lanturi polinucleotidice, orientate cu bazele spre interiorul structurii, astfel incat fata in fata sa se gaseasca intotdeauna fie adenina si timina, fie citozina si guanina. Structura spatiala astfel formata se stabilizeaza prin doua legaturi de hidrogen intre timina si adenina si trei legaturi de hidrogen intre citozina si guanina.
Functia nucleului bacterian consta in depozitarea informatiei genetice necesara autoreplicarii precum si pentru organizarea structurala si functionala a celulei bacteriene. Constituie sediul ereditatii cromozomiale si asigura toate caracterele specifice de specie ale bacteriei respective.
- Autoreplicarea ADN (diviziunea nucleului) precede diviziunea celulara si este de tip semiconservativ, adica fiecare molecula de ADN nou formata contine un lant polinucleotidic din molecula de ADN parentala si un lant polinucleotidic nou sintetizat. Nucleul are rol esential in multiplicarea bacteriilor. Diviziunea nucleului incepe printr-un clivaj longitudinal al cromozomului, catalizat de ADN-polimeraza si urmat apoi de resinteza catenei complementare. In celula apar doua lanturi bicatenare (identice intre ele si identice cu parentalul) care se separa si migreaza spre polii celulei (impreuna cu mezozomii care s-au dedublat concomitent cu nucleul). Apare peretele despartitor, iar cele doua celule se separa. In acest fel se transmit toate caracterele de specie la descendenti.
- Heteroreplicarea (sinteza proteinelor proprii bacteriei). Nucleul dirijeaza aceasta sinteza pe baza informatiei continuta in ADN. Informatia genetica este transmisa de la molecula de ADN la ribozomi prin intermediul ARN mesager (ARNm) sau informational (prima transcriptie). Sinteza de ARNm (lant unic de nucleotide, fiecare nucleotid avand in structura riboza, o baza azotata - guanina, citozina, adenina sau uracil- si o molecula de acid fosforic) este catalizata de ARN-polimeraza pe modelul constituit de unul din lanturile ADN. Moleculele de ARNm migreaza apoi in citoplasma la sediile de sinteza a proteinelor reprezentate de ribozomi, unde vor servi ca tipar sau matrita pentru asamblarea acizilor aminati in lanturi polipeptidice (a doua transcriptie). Aminoacizii de structura activati enzimatic sunt transportati din citoplasma la ribozomi de molecule de ARN de transport sau solubil (ARNt) specifice fiecarui aminoacid.
In afara de ADN cromozomial, la unele bacterii sunt prezente molecule circulare mici, extracromozomiale de ADN care se numesc plasmide si care se replica independent de cromozomul bacterian.
2.2.2. CITOPLASMA
Situata intre materialul nuclear si fata interna a membranei citoplasmatice, citoplasma este un sistem coloidal complex alcatuit din aproximativ 80% apa, in care se gaseste o cantitate mare de molecule organice mici (rezultate ale metabolismului bacterian), ioni anorganici, enzime si acizi ribonucleici (ARN ribozomal, ARN de transport, ARN mesager). In functie de specie, in citoplasma bacteriilor se mai pot gasi: plasmide, vacuole si incluzii. Este netimentata fiind lipsita de unele organite celulare prezente la celulele eucariote, cum sunt reticulul endoplasmatic, aparatul Golgi, mitocondriile.
a. Ribozomii
Principalele elemente ale citoplasmei, sunt structuri sferice, mai mici decat ribozomii celulelor eucariote, si reprezinta sediul sintezelor proteice din celula.
Au constanta de sedimentare de 70S (completi), ce se mentine stabila in prezenta unei anumite concentratii de Mg2+ si K+ din citoplasma. In absenta ionilor de Mg2+ are loc disocierea ribozomilor in subunitati de 50S si 30S (incompleti), subunitati care reprezinta tinta pentru actiunea unor antibiotice (streptomicina, eritromicina, cloramfenicol).
Din punct de vedere chimic sunt alcatuiti din 60% ARN si 40% proteine. O parte de ribozomi se asociaza formand polizomi (mai ales in timpul sintezelor proteice), altii sunt liberi, iar o a treia categorie se ataseaza mezozomilor sau membranei citoplasmatice.
La nivelul ribozomilor sunt sintetizate toate enzimele necesare metabolismului care caracterizeaza fiziologia bacteriei.
b. Mezozomii
Sunt structuri membranare care se formeaza prin invaginarea membranei citoplasmatice sub forma de buzunar sau in deget de manusa, prezente la bacteriile Gram pozitive si ocazional la cele Gram negative. Ei formeaza in citoplasma cavitati deschise spre spatiul periplasmic (spatiul dintre membrana citoplasmatica si peretele celular) si sunt in contact direct cu materialul nuclear. Au o organizare mai complexa la bacteriile Gram pozitive si sunt mai rudimentari la bacteriile Gram negative.
Functiile mezozomilor
participa la replicarea cromozomului bacterian si diviziunea celulara;
sediul unor enzime hidrolitice care indeplinesc rolul enzimelor lizozomale de la celulele eucariote;
sinteza si secretia unor exoenzime (penicilinaza sau cefalosporinaza);
participa la reactiile de fosforilare oxidativa si oxidoreducere (minor).
c. Plasmidele
Organite specifice celulei procariote, reprezinta unitati genetice extracromozomiale prezente numai la unii indivizi bacterieni (se pot transfera de la un individ bacterian la altul). Sunt molecule circulare de ADN, mici, capabile sa se replice independent de cromozomul bacterian si care sunt responsabile de ereditatea extracromozomiala (rol in transmiterea unor caractere cum este rezistenta la antibiotice).
d. Incluziile granulare (depozite de substante de rezerva)
Descrise la unele specii bacteriene, sunt formatiuni structurale inerte, temporare, de diferite dimensiuni, variind in functie de specia bacteriana si conditiile de mediu.
Compozitia lor chimica este diferita, ele putand fi de:
glicogen (la specii din familia Enterobacteriaceae);
asemanatoare amidonului (la unii bacili aerobi sporulati - genul Clostridium);
lipide (la genul Bacillus);
polimetafosfati (sau incluziile de volutina descrise de Babes si Ernst la bacilii difterici), etc.
Incluziile sunt structuri legate de activitatea metabolica a celulei bacteriene si reprezinta un material de rezerva care poate fi folosit ca sursa de energie.
e. Vacuolele
Vacuolele, alte organite intracitoplasmatice, sunt formatiuni sferice, care contin substante lichide sau gazoase, inconjurate de un invelis lipoproteic unistratificat.
f. Oxizomii
Reprezinta sediul enzimelor de oxidoreducere. Sunt organite specifice celulei procariote. In oxizomi se gasesc citocromii, citocromoxidaza, flavin-enzima etc., iar in oxizomi si materialul solubil se gasesc si enzimele ciclului Krebs: SDH (se gaseste in cantitate crescuta in oxizomi si in cantitate scazuta in materialul solubil); malic-DH (se gaseste in cantitati egale, atat in oxizomi, cat si in materialul solubil); aconitaza, izocitric-DH, a-Ketoglutaric-DH (se gasesc in cantitati scazute in oxizomi si in cantitati crescute in materialul solubil).
Apare ca un strat foarte subtire care inconjoara citoplasma si este foarte aderenta de peretele celular. Este o membrana fina (6,5-7nm), elastica, lipsita de rezistenta mecanica, reprezentand zona unde se produc fenomenele de permeabilitate osmotica si selectiva (membrana semipermeabila).
Pe sectiune, membrana apare trilaminata, avand drept compozitie chimica doua straturi fosfolipidice dispuse cu partile hidrofobe fata in fata. Printre moleculele fosfolipidice se gasesc molecule proteice (grupari polare, permeaze-translocaze), situate fie la nivelul unuia dintre cele doua straturi fosfolipidice, fie le traverseaza fiind expuse la ambele fete ale membranei.
Functional, membrana indeplineste urmatoarele roluri:
este o membrana semipermeabila, care regleaza schimburile ce au loc intre celula bacteriana si mediul extern, atat prin procese active specifice, cat si prin procese de difuziune pasiva;
secreta numeroase enzime hidrolitice ce se elibereaza in mediul inconjurator unde scindeaza macromoleculele in molecule mai mici, ce pot fi transportate in interiorul celulei;
participare la sisteme chemotactice: la nivelul membranei sunt prezenti receptori asupra carora actioneaza stimulii chimici din mediu cu rol de atractie (atractanti) sau de respingere (repelenti);
indeplineste functia bioenergetica, de eliberare a energiei prin fosforilare oxidativa. In membrana este structurat lantul respirator si unele enzime din ciclul Krebs (dehidrogenaze), membrana procariota preluand functia mitocondriilor din celula eucariota;
participa activ la cresterea si diviziunea celulei bacteriene, la formarea sporului bacterian;
reprezinta structura celulara tinta pentru detergenti care, utilizati ca substante dezinfectante altereaza structura acesteia, sau pentru unele antibiotice care interfereaza cu functia biosintetica a membranei (polimixinele).
Peretele celular este o structura unica pentru bacterii, responsabila de forma si rigiditatea celulei (participa minor la osmoza), localizata la exteriorul membranei citoplasmatice si prezenta la toate bacteriile, cu exceptia reprezentantilor genului Mycoplasma si Archaebacteriilor. Peretele este format dintr-un strat bazal, asemanator la toate bacteriile si un strat al structurilor superficiale, foarte diferentiat, in functie de care bacteriile manifesta caractere tinctoriale diferite: bacterii Gram pozitive, Gram negative si acido-alcoolorezistente.
Reteaua de peptidoglican (mureina) este structura chimica responsabila de rigiditatea peretelui celular si care asigura forma si rezistenta mecanica a bacteriei. Reteaua de peptidoglican, prezenta la toate bacteriile, este formata din 3 portiuni (ura 5):
- Scheletul de baza alcatuit din molecule lungi paralele polizaharidice de N-acetil-glucozamina (N-Ac-Glc) si acid N-acetil-muramic (N-Ac-Mur);
![]() D-ALA
D-ALA
N-Ac-Glc N-Ac-Mur N-Ac-Glc N-Ac-Mur
L-ALA (GLY) 5
D-GLU
L-LYS
![]() D-ALA
D-ALA
N-Ac-Glc N-Ac-Mur N-Ac-Glc N-Ac-Mur
L-ALA (GLY) 5
D-GLU
L-LYS
![]() D-ALA
D-ALA
(GLY) 5
ura 5: Reteaua de peptidoglican (dupa C. Voiculescu, 1996)
- Componenta peptidica formata din punti tetrapeptidice identice, transversale, intre unitatile de N-Ac-Mur a doua lanturi vecine. Structura acestor punti difera la bacteriile Gram pozitive de cele Gram negative si chiar de la specie la specie. In aceasta structura intra D si L aminoacizi: L-alanina, D-glutamina, L-lizina (sau diaminopimelic la bacteriile Gram negative), D-alanina. Aceasta componenta se sintetizeaza initial ca un pentapeptid, dar intr-un peptidoglican complet se gaseste sub forma de tetrapeptid deoarece ultimul aminoacid, D-alanina, este inlaturat cand un lant peptidic se leaga de un altul vecin;
- Punti incrucisate de pentaglicina (intre pozitia 4 si 3 a doua punti tetrapeptidice vecine) la bacteriile Gram pozitive. La bacteriile Gram negative, tetrapeptidele se leaga intre ele printr-o simpla legatura peptidica.
Structura stratului structurilor speciale
Se deosebesc trei tipuri de structuri speciale: Gram pozitiv, Gram negativ si acido-alcoolorezistent.
a. Bacterii Gram pozitive (caracteristici particulare)
Peretele celular al bacteriilor Gram pozitive este relativ mai gros, dar cu o compozitie mai simpla. Peptidoglicanul reprezinta 50-90% din greutatea uscata a peretelui celular, are o grosime de 15-30 nm si contine pana la 200 de lanturi paralele de mureina, legate tridimensional pentru a forma o retea groasa. Stratul structurilor speciale este redus si alcatuit din polimeri hidrosolubili care sunt acizii theicoici: acidul ribitoltheicoic si gliceroltheicoic. Acestia pot patrunde pana la membrana citoplasmatica (acizii theicoici cu glicerol) legandu-se covalent de aceasta - acizii theicoici de membrana sau numai pana la perete - acizii theicoici de perete (acizi theicoici cu ribitol). Acizii theicoici intervin in special in cresterea si diviziunea celulei. Ei reprezinta determinanti antigenici majori care stau la baza identificarii serologice a unor bacterii, fiind substante care, patrunse in organism, induc un raspuns specific din partea sistemului imun al organismului.
De asemenea, la unele specii, se evidentiaza si prezenta de polizaharide: polimeri de manoza, arabinoza, galactoza, glucozamina si polimeri de zaharuri acide (acid glucuronic, acid manuronic).
Peretele celular al bacteriilor Gram pozitive este sensibil la actiunea lizozimului (enzima hidrolitica prezenta in lacrimi, mucus, saliva) care rupe legaturile dintre acidul N-acetil muramic si N-acetilglucozamina, autolizinelor bacteriene (enzime muralitice) care intervin in diviziunea celulei bacteriene, penicilinei care inhiba sinteza peptidoglicanului.
b. Bacterii Gram negative (caracteristici particulare)
Peretele este mai subtire dar mult mai complex structurat, multistratificat. Peptidoglicanul are o grosime de 4-5 nm, reprezentand numai 10% din greutatea uscata a bacteriei. Stratul superficial este insa mult mai complex decat la bacteriile Gram pozitive, alcatuit din spatiul periplasmic, membrana externa si lipopolizaharidul de perete (LPS).
Principalele diferente structurale ale peretelui celular bacterian la bacteriile Gram pozitive si Gram negative sunt prezentate in ura 6.
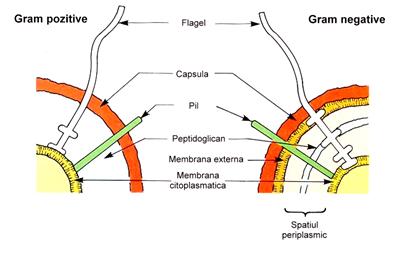
ura 6: Diferentele structurale ale peretelui celular la bacteriile Gram pozitive si Gram negative (dupa Murray, Drew, Kobayashi, Thompson, 1990)
Este un timent cu functie de stocare ce se intinde de la membrana citoplasmatica pana la membrana externa. Largimea spatiului periplasmic este variabila, depinzand de: starea fiziologica a celulei; conditiile de cultivare.
El contine peptidoglicanul si un gel care favorizeaza nutritia bacteriei prin continutul in enzime degradative (fosfataze, nucleaze, proteaze). Tot aici sunt prezente enzimele de inactivare ale unor antibiotice (betalactamaze, cefalosporinaze). Enzimele stocate la nivelul spatiului periplasmic sunt eliberate prin membrana externa in functie de necesitati si aceasta este una din explicatiile marii capacitati de adaptare la mediu a bacteriilor Gram negative spre deosebire de cele Gram pozitive care, neavand membrana externa, elibereaza enzimele imediat ce sunt sintetizate.
Este asemanatoare ca structura cu membrana citoplasmatica fiind formata dintr-un strat dublu de fosfolipide in care sunt inclavate proteine de o diversitate foarte mare: unele dintre acestea strabat membrana si pot ajunge pana la peptidoglican, altele sunt atasate pe fata interna sau externa a membranei si proemina in spatiu sau la suprafata, iar altele sunt libere (urile 7 si 8).
In membrana externa exista proteine majore (porine, non porine) si proteine minore. Porinele sunt cele care penetreaza ambele fete ale membranei externe si formeaza pori sau canalicule, prin care difuzeaza spre interiorul celulei molecule cu greutate moleculara pana la 600 D. Substantele nutritive ajung din exteriorul celulei pana la spatiul periplasmic, unde se petrec evenimente metabolice de degradare in substante ce pot fi transportate prin membrana citoplasmatica in interiorul celulei. A doua categorie de proteine majore, non porinele, sunt fie receptori pentru pili sexuali, fie enzime care regleaza sinteza capsulei bacteriene. Proteinele minore functioneaza ca transportori transmembranari specifici pentru moleculele mici (ionii Fe3+, vitamine).
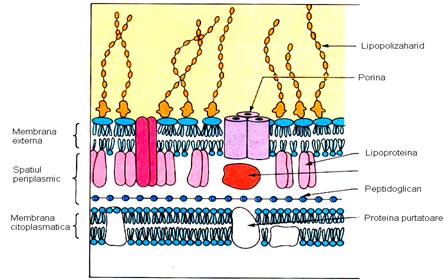
ura 7: Peretele celular la bacteriile Gram negative (dupa Murray, Drew, Kobayashi, Thompson 1990)
Lipopolizaharidul de perete (LPS)
Deasupra membranei externe a bacililor Gram negativi se afla lipopolizaharidul de perete (LPS) sau endotoxina bacililor Gram negativi. LPS este o toxina termolabila care se elibereaza in mediul inconjurator numai dupa liza acestor bacterii, fiind foarte reactiva in organismul gazda.
In structura lipopolizaharidului intra: lipidul A, miezul sau core si unitati monozaharidice repetitive.
lipidul A cu o structura particulara, responsabil de toxicitate: produce febra, activeaza mecanismele apararii antiinfectioase si, in exces, produce socul endotoxic cu evolutie grava, chiar fatala;
miezul sau core (polizaharid), numit si antigen R, comun tuturor bacteriilor Gram negative;
unitati monozaharidice repetitive (15-40) care sunt specifice de specie si tip si constituie antigenul O al bacteriilor Gram negative (ura 8).
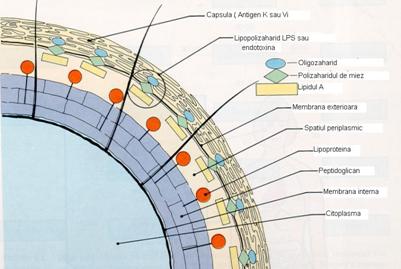
ura 8: Peretele celular la bacteriile Gram negative (dupa Murray, Drew, Kobayashi, Thompson, 1990)
c. Bacteriile acido-alcoolo rezistente
Dintre aceste bacterii, bacilul tuberculos si bacilul leprei, reprezentanti ai genului Mycobacterium, sunt de interes medical. Aceste bacterii au peretele celular asemanator cu cel al bacteriilor Gram pozitive. Structurile speciale contin lipide pana la 30%, aproape jumatate din acestea fiind reprezentate de acidul micolic si o ceara ce confera acestor bacterii caractere tinctoriale deosebite si rezistenta crescuta la factorii de mediu. Astfel daca, dupa o incalzire de scurta durata ce topeste cerurile, un colorant patrunde in celula bacteriana, decolorarea acesteia sub actiunea acizilor sau alcoolilor nu se mai produce ca la alte bacterii. Acest caracter se numeste acido-alcoolorezistenta. Aceste bacterii se coloreaza foarte slab in coloratia Gram, evidentierea lor facandu-se la cald prin tehnica Ziehl-Neelsen.
d. Bacterii cu perete alterat sau formele L
Sunt bacterii cu stratul bazal viciat sub actiunea unor factori din mediu ca, de exemplu, lizozimul, care lizeaza peptidoglicanul, si penicilina care impiedica sinteza acestuia.
e. Bacterii fara perete celular
Exista bacterii lipsite in mod natural de perete celular, care nu pot avea o forma constanta, forma lor fiind variabila in functie de mediul in care se afla. Exemplu: Mycoplasma (cu forma cocoidala, de para, alungita sau filamentoasa).
Functiile peretelui celular
asigura forma, rezistenta mecanica si osmotica a bacteriei
participa la diviziunea celulei si in cresterea acesteia, urmand membrana citoplasmatica in formarea septurilor transversale care separa celula mama in celule fiice; protoplastii si sferoplastii, lipsite de perete celular, nu se divid;
stocheaza unele enzime in spatiul periplasmic la bacteriile Gram negative ce vor fi eliberate dupa necesitati;
prezinta receptori pentru bacteriofagi (virusuri care paraziteaza bacteriile) prin proteinele de membrana externa, initiind infectia virala a celulei bacteriene;
este sediul antigenelor de suprafata, fiind deci implicat in raspunsul imun al macroorganismului;
este sediul unor factori de patogenitate
are rol in procesul de sporulare;
reprezinta tinta de actiune pentru unele antibiotice si detergenti;
prin structura diferita separa cele doua categorii de bacterii in coloratia Gram (Gram pozitive si Gram negative) sau confera caracter de acidoalcoolorezistenta.
Zona externa sau cortexul ovarului contine celule germinative (Figura 9-1). Aceste celule germinative se gasesc in numerosi foliculi ovarieni. St [...] |
DIAGNOSTICUL DE LABORATOR AL INFECTIILOR PRODUSE DE STREPTOCOCI Manifestarile clinice determinate de streptococi includ: - infectii acute sup [...] |
FACTORII DE REZISTENTA A ORGANISMULUI INFECTAT Organismul este expus in toate etapele vietii la diferite infectii, cea mai mare receptivitate [...] |
Copyright © 2010 - 2024
: eSanatos.com - Reproducerea, chiar si partiala, a materialelor de pe acest site este interzisa!
Informatiile medicale au scop informativ si educational. Ele nu pot inlocui consultul medicului si nici diagnosticul stabilit in urma investigatiilor si analizelor medicale la un medic specialist.
Termeni si conditii - Confidentialitatea datelor - Contact