Intre virusul animal si celula gazda pe care o paraziteaza se pot stabili doua feluri de relatii: de tip litic (citocid) si de tip simbiotic.
1. Relatii virus-celula gazda de tip litic (citocid)
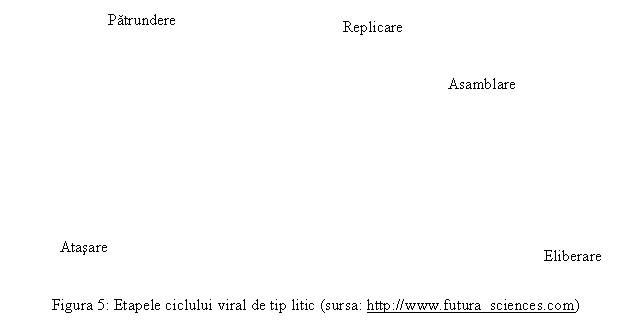
In acest tip de relatie (urile 4 si 5), infectia cu virus a unei celule animale se soldeaza cu moartea celulei si cu eliberarea de particule de virus neoformate (progeni) in mediul extracelular.
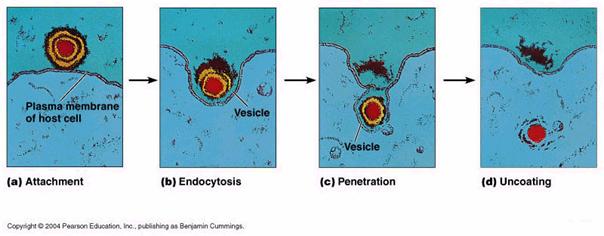
Etapele acestei relatii sunt: adsorbtia, penetrarea, decapsidarea, sinteza particulelor virale neoformate, eliberarea particulelor virale neoformate.
Adsorbtia
Procesul implica doi timpi: colizinea si atasarea.
Coliziunea reprezinta alipirea particulei (prin situsurile virale de atasare) de suprafata celulei sensibile (tabel 2). Ea depinde, pe de o parte, de proportia intre numarul de virioni si numarul de celule (raportul de infectivitate), iar pe de alta parte, de unele conditii ale mediului de reactie (temperatura, pH, concentratie ionica).
Atasarea semnifica unirea stransa intre virion si unele zone de pe suprafata celulei, numite receptori.
Receptorii celulari pot fi specifici anumitor agenti virali, explicand tropismul unor virusuri fata de anumite substrate celulare (myxovirusuri, picornavirusuri) sau pot fi nespecifici (neselectivi in raport cu mai multe categorii de virusuri - arbovirusuri, poxvirusuri).
Atasarea specifica a myxovirusurilor (v. gripale) si paramyxovirusurilor (v. urlian, v. bolii Newcastle) se datoreste interventiei unei enzime virale (neuraminidaza) care prezinta proprietatea de a ataca substanta receptor constituita din situsuri de mucoproteina (bogate in acid neuraminic sau sialic).
|
Agent viral |
Situs viral de atasare |
Receptori celulari |
|
Virus gripal |
hemaglutinina, neuraminidaza (invelis) |
mucoproteine (acid sialic) |
|
Virus HIV |
gp 120 (invelis) |
receptor limfocitar CD4 |
|
Virus poliomielitic |
situs capsidal |
receptor specific (cu exprimare diferita in vivo sau in vitro pe celula renala de maimuta) |
|
Virus rabic |
hemaglutinina (invelis) |
Receptor acetilcolinic (muschi striat) |
|
Virus Herpes simplex |
gb (invelis) |
Receptor pe celula fibroblastica de hamster, soarece, om |
Tabel 2: Exemple de situsuri virale de atasare si receptori celulari
Atasarea specifica a picornavirusurilor a fost cercetata indeosebi la poliovirusuri. Astfel, extragandu-se un material proteic din membranele celulelor susceptibile la infectia cu poliovirusuri, s-a demonstrat capacitatea acestui material de a inactiva virusul. In prima etapa a atasarii, receptorii specifici poliovirusurilor realizeaza un complex reversibil, pentru ca, in timp, legatura receptor-virus sa devina stabila (capacitatea infectiva a virusului nu mai poate fi recuperata prin diverse tentative de desfacere a complexului). Tot referitor la receptorii pentru poliovirusuri este de remarcat ca, uneori, celulele apartinand unei anumite specii sau unui anumit allotip, rezistente in vivo la infectia poliovirotica, devin deosebit de sensibile la acelasi virus, odata cu scoaterea lor din organism si cultivarea in vitro (exemplu: celulele renale de maimuta). Acest fenomen poate fi explicat prin derepresarea in vitro a unei gene celulare care codifica exprimarea activitatii receptorului de membrana pentru poliovirus. Pe de alta parte, sunt si situatii in care poliovirusul se ataseaza pe receptorii celulari, fara a se declansa etapele consecutive ale infectiei de tip litic, ceea ce releva complexitatea fenomenului de atasare, in functie de tipul de celula si de virusul infectant.
Penetrarea
In aceasta etapa are loc trecerea virusului din exteriorul celulei in interiorul acesteia. Inglobarea virusului in citoplasma (viropexie) a fost confirmata numai in unele cazuri de relatie de tip litic (poliovirusuri, myxovirusuri, reovirusuri), prin prezenta in citoplasma celulelor virus infectate, in primele ore de la initierea infectiei virale, a unor vacuole de fagocitare care contin particule virale intacte.
Desfacerea capsidei
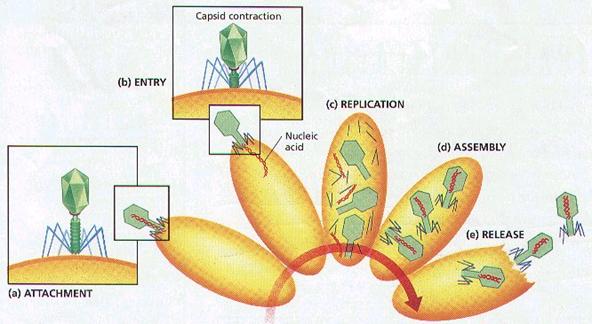
Spre deosebire de bacteriofagi, la care numai acidul nucleic este injectat in celula parazitara, la virusurile animale se impune pierderea unei parti a capsidei virale, prin enzimele lizozomale ale celulei. Lizozomii se aliniaza de-a lungul peretilor vacuolelor de fagocitare, eliberand enzime litice care desfac atat peretele vacuolei, cat si o parte a capsidei virionilor continuti in vacuola.
Pentru virusurile ADN mari (poxvirusuri) decapsidarea este catalizata si de enzime virus-induse in celula gazda.
Sinteza particulelor virale neoformate
Este cea mai importanta etapa a relatiei virus celula gazda de tip litic si consta in esenta, din codificarea unui nou program, folosind aparatul ribozomal al celulei parazitate. Aceasta etapa cuprinde doua categorii de fenomene: precoce (early) si tardive (late).
In primele 6-8 ore de la patrunderea virusului in celula, acidul nucleic viral induce doua procese:
- Oprirea sintezelor proteice de tip celular, prin paralizarea functionala a ADN celular;
Codificarea unor enzime numite polimeraze virus-specifice (ARN polimeraza virus-indusa, in cazul virusurilor ARN; ADN polimeraza virus-indusa in cazul virusurilor ADN). Aceste polimeraze catalizeaza formarea de copii ale acidului nucleic viral, folosind ca matrice nucleoidul viral decapsidat dupa intrarea in celula. Se initiaza, in acest fel, replicarea acidului nucleic viral, cu aparitia unor multiple copii continand genele de tip viral.

![]()
Copiile de acid nucleic viral programeaza la nivelul ribozomilor celulari sinteze proteice noi, folosind ca materie prima aminoacizii celulari, prin legarea acestora intr-o ordine (secventa) diferita, conform modelului inscris in genomul viral.
In cazul virusurilor ARN, moleculele de acid nucleic viral rezultate din replicarea intracelulara reprezinta ele insele un ARN mesager nou, citit direct de ribozom.
In cadrul virusurilor ADN, pe matricea acidului nucleic viral se sintetizeaza mai intai un ARN mesager virus specific, care functioneaza apoi ca un nou programator la nivelul ribozomilor.
- Proteinele virale sintetizate de catre ribozomi, potrivit unui nou program, se aseaza in jurul moleculelor de acid nucleic viral si astfel se produce asamblarea si maturarea particulelor virale neoformate. In cadrul acestui proces, are loc si asezarea subunitatilor capsidale (monomerilor) dupa simetria caracteristica tipului de virus infectant, fenomen codificat si el de catre unele gene virale.
Eliberarea particulelor virale neoformate
Virionii nou formati, care, cu rare exceptii (mutante) sunt identici morfologic si genetic cu particula care a initiat infectia virala a celulei, sunt eliberati in mediul extracelular. De regula, acest fenomen se insoteste de dezintegrarea (moartea) celulei gazda.
In urma acestui tip de relatie virus-celula gazda, celula nu moare, ci castiga proprietati noi.
Etapele infectiei virale de tip simbiotic sunt asemanatoare cu cele din infectia de tip litic in ce priveste adsorbtia, penetrarea si desfacerea capsidei. Deosebirile apar in etapele urmatoare: programarea sintezelor de tip viral, aparitia unor proprietati de tip nou.
Inducerea unui nou program de catre acidul nucleic viral in celula animala virus infectata nu se soldeaza cu oprirea activitatii ADN celular, astfel incat ribozomii vor sintetiza concomitent proteine celulare si virale. Modul programarii sintezelor virale difera la virusurile ADN si ARN capabile sa stabileasca relatie simbiotica.
Virusurile ADN
ADN viral patruns in celula poate urma doua modele de codificare a sintezelor proprii (urile 6 si 7).
Modelul 1: ADN viral se integreaza (insera) intr-o zona a ADN celular. Ulterior, se va sintetiza un ARN mesager complementar mixt, continand anticodoni de tip celular si anticodoni de tip viral. ARN mesager va ajunge la ribozomi, care vor citi mesajul.
Model 2: ADN viral nu se integreaza in genomul celular, ci induce sinteza unei enzime precoce (ARN-polimeraza virus indusa) care catalizeaza sinteza de ARN mesager nou, pe matricea de ADN viral. Pe de alta parte, ADN celular comanda sinteza de tip celular prin intermediul ARN mesager normal. In acest fel, unii ribozomi vor citi mesajul de pe molecula de ARN mesager celular, altii de pe ARN mesager de tip viral, in aceeasi celula aparand proteine celulare si virale.
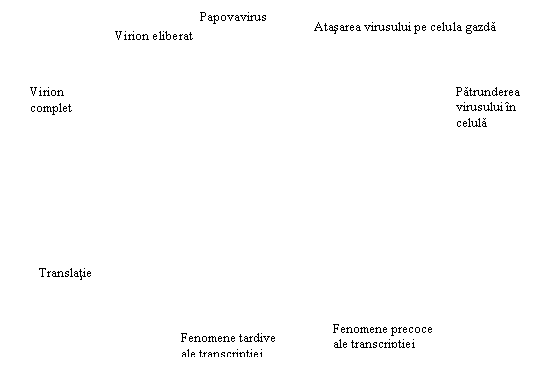
ARNm Proteine capsidale
![]()
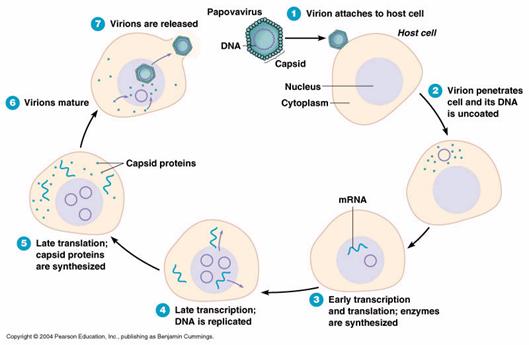
ura 6: Ciclul
replicativ viral (sursa: https://www.futura_sciences.com)
Astfel, un acelasi ribozom va sintetiza proteine cu secvente de aminoacizi dictate de ADN celular, precum si proteine cu secvente de aminoacizi dictate de ADN viral.
Virusurile ARN
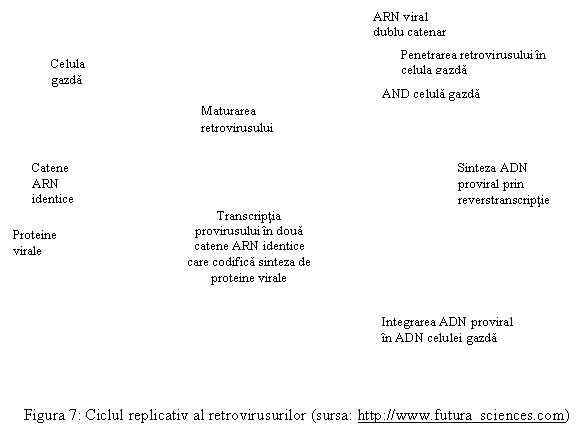
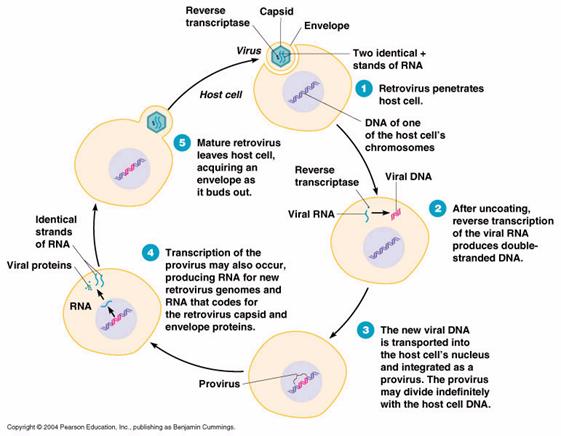
In cazul virusurilor ARN, mecanismul integrarii acidului viral in ADN celular a fost propus de Temin 1972, modelul fiind confirmat apoi experimental la virusurile integrate de tip C (retrovirusuri). Procesul este cunoscut sub numele de revers-transcriptie. ARN viral patruns in celula reprezinta o noua matrice, care induce formarea unei enzime, numita revers-transcriptaza. Aceasta enzima catalizeaza sinteza unui ADN complementar folosind ca matrice ARN viral (invers decat la celula eucariota normala, unde pe matricea de ADN se formeaza ARN complementar).
ADN complementar viral (provirus) se integreaza in ADN celular si apoi urmeaza aceeasi cale ca la modelul 1 de programare de la virusurile simbiotice ADN, rezultand proteine mixte celular-virale.
Revers-transcriptia, caracteristica pentru virusurile oncogene tip C si pentru virusul HIV, implica interventia mai multor enzime, dupa cum urmeaza (tabel 3):
|
Secventa a revers-transcriptiei |
Enzime virale responsabile |
|
Formarea primei catene de ADN proviral |
Revers-transcriptaza |
|
Depolimerizarea nucleotidelor din ARN viral care nu functioneaza ca matrice |
Ribonucleaza H |
|
Formarea ADN proviral dublu catenar (atasarea celei de a doua catene) |
Revers-transcriptaza |
|
Integrarea provirusului in ADN celular |
Integraza |
|
Activarea genomului viral activat |
Complex genic de activare (exemplu: tat/nef la virusul HIV), plus un complex enzimatic |
Tabel 3: Etape ale revers-transcriptiei si enzimele responsabile
![]()
Aparitia unor proprietati noi
Consecinta stabilirii unei relatii de tip simbiotic virus-celula gazda, consta in aparitia unor proprietati modificate ale celulei infectate fata de celula normala-neinfectata.
Printre proprietatile modificate mentionam:
Transformarea celulara: implica modificari profunde morfologice, genetice, biochimice, de suprafata, de potential oncogen, proces complex, echivalent pana la un punct malignizarii;
Exprimarea de antigene noi pe suprafata celulei
Instalarea unor modele metabolice particulare, care asigura un echilibru intre sinteza concomitenta de proteine de tip celular si de tip viral in aceeasi celula.
Mecanismele care stau la baza aparitiei acestor noi proprietati sunt, pe de o parte, introducerea, concomitent cu acidul nucleic viral, a unor gene noi, capabile sa codifice proprietatile modificate (exemplu: genele pol, gag, env, onc, ale virusurilor oncogene ARN-retrovirusuri, codifica sinteza revers-transcriptazei, proteinelor p10, p12, p15, p30, glicoproteinei majore gp70, malignizarea celulei), iar pe de alta parte, derepresia unor gene celulare datorita patrunderii si integrarii acidului nucleic viral in genomul celulei cu codificarea consecutiva a exprimarii de noi proprietati (exemplu: antigenul carcino-embrionar, prezent numai in celulele embrionate si fetale si, respectiv, in celula cancerizata, dar absent in celula normala adulta).
Intre virus si celula-gazda, exista posibilitatea coexistentei celor doua tipuri de relatii: litica si simbiotica (virusurile de tip C la multe specii de mamifere). In acest caz, intr-o populatie celulara sau intr-un tesut cancerizat, unii virioni induc transformare (prin integrarea acidului nucleic viral in genomul celular), altii induc relatie litica (producerea de progeni virali, eliberarea de particule neoformate si moartea celulei gazda).
O alt caz deosebit este oferit de virusurile lente. Acesti agenti virali stabilesc o relatie de tip litic, cu o rata foarte joasa de infectivitate, astfel incat, in acelasi tesut, unele celule prezinta leziuni, altele sunt normale. Consecinta este evolutia lunga (ani, decenii) a unei afectiuni degenerative, la care se poate adauga apoi si un mecanism autoimun de intretinere.
Un alt aspect particular al relatiilor virus-celula gazda este reprezentat de virusurile mascate (virusuri din grupul Herpes, v. rubeolic, v. rabic, v. HIV). Aceste virusuri, in anumite circumstante, pot produce o infectie latenta a celulei, genomul viral fiind integrat in genomul celular fara sa-si tradeze prezenta. La un moment dat, la interventia unui factor demascator, genomul viral intra in activitate, se desprinde din insertia cu genomul celular si declanseaza ciclul de multiplicare virala de tip litic, cu dezintegrarea celulei gazda.
Sarcina, travaliul si nasterea pot afecta pentru un timp relatiile sexuale in cazul multora dintre cupluri. Spre sfarsitul sarcinii, raporturile sexua [...] |
Sarcina, travaliul si nasterea pot afecta pentru un timp relatiile sexuale in cazul multora dintre cupluri. Spre sfarsitul sarcinii, raporturile sexua [...] |
Virusurile, ca si alti agenti infectiosi nu sunt antigene inerte. Virusurile evolueaza pe masura ce organismul gazda elaboreaza mecanisme nespecifice [...] |
Copyright © 2010 - 2024
: eSanatos.com - Reproducerea, chiar si partiala, a materialelor de pe acest site este interzisa!
Informatiile medicale au scop informativ si educational. Ele nu pot inlocui consultul medicului si nici diagnosticul stabilit in urma investigatiilor si analizelor medicale la un medic specialist.
Termeni si conditii - Confidentialitatea datelor - Contact